Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out Of Africa Fulvio Cruciani,
We explored the phylogeography of human Y-chromosomal haplogroup E3b by analyzing 3,401 individuals from five continents. Our data refine the phylogeny of the entire haplogroup, which appears as a collection of lineages with very different evolutionary histories, and reveal signatures of several distinct processes of migrations and/or recurrent gene flow that occurred in Africa and western Eurasia over the past 25,000 years. In Europe, the overall frequency pattern of haplogroup E-M78 does not support the hypothesis of a uniform spread of people from a single parental Near Eastern population. The distribution of E-M81 chromosomes in Africa closely matches the present area of distribution of Berber-speaking populations on the continent, suggesting a close haplogroupethnic group parallelism. E-M34 chromosomes were more likely introduced in Ethiopia from the Near East. In conclusion, the present study shows that earlier work based on fewer Y-chromosome markers led to rather simple historical interpretations and highlights the fact that many population-genetic analyses are not robust to a poorly resolved phylogeny.
The human Y-chromosome haplogroup E is characterized by the mutations SRY4064, M96, and P29, on a back- ground defined by the insertion of an Alu element (YAP ) (Y Chromosome Consortium 2002; Jobling and Tyler- Smith 2003). Two of the three branches of haplogroup E, the major clades E1 and E2, have been observed almost exclusively on the African continent, where their distri- bution has been analyzed in detail (Underhill et al. 2000; Cruciani et al. 2002). The third branch, the clade E3, defined by the mutation P2, is the only one that has also been observed in Europe and in western Asia, where it has generally been found at frequencies !25% (Hammer et al. 2000, 2001; Semino et al. 2000; Scozzari et al. 2001; Cinniog˘lu et al. 2004). On the basis of the previously published phylogeny (Y Chromosome Consortium 2002; Jobling and Tyler- Smith 2003), the mutations M2/P1/M180, on the one hand, and M35/M215, on the other, further subdivide E3 in two monophyletic haplogroups: E3a and E3b. Both haplogroups are frequent in Africa (Underhill et al. 2000; Cruciani et al. 2002), although, to date, only E3b has also been observed in Europe (Semino et al. 2000) and western Asia (Underhill et al. 2000; Cinniog˘lu et al. 2004). Recently, it has been proposed that E3b originated in sub-Saharan Africa and expanded into the Near East and northern Africa at the end of the Pleis- tocene (Underhill et al. 2001). E3b lineages would have then been introduced from the Near East into southern Europe by immigrant farmers, during the Neolithic ex- pansion (Hammer et al. 1998; Semino et al. 2000; Un- derhill et al. 2001). The three main subclades of haplogroup E3b (E-M78, E-M81, and E-M34) and the paragroup E-M35* are not homogeneously distributed on the African continent: E- M78 has been observed in both northern and eastern Africa, E-M81 is restricted to northern Africa, E-M34 is common only in eastern Africa, and E-M35* is shared by eastern and southern Africans (Cruciani et al. 2002). Given the strong geographic structuring observed for the four subsets of E3b within Africa, it is possible that different E3b lineages also have different frequency pro- files in western Eurasia and that the evolutionary events underlying the introduction of E3b chromosomes in this area from Africa were not as simple (Rosser et al. 2000; Richards et al. 2002; Jobling and Tyler-Smith 2003) as previously proposed (Hammer et al. 1998; Semino et al. 2000; Underhill et al. 2001). In the present study, we address the question of the origin and dispersal of haplogroup E3b subclades within and outside of Africa by analyzing 3,401 individuals from five continents. These include 1,510 individuals analyzed here for the first time for Y-chromosome mark- ers (see also footnotes b, c, and d of table 1). All of the subjects were typed for the YAP polymor- phism (Hammer and Horai 1995), and those who were YAP (haplogroup DE) were analyzed for the SRY4064 (Whitfield et al. 1995), M35, and M215 mutations (Un- derhill et al. 2000, 2001). Two subjects were found to carry the derived state at M215 and the ancestral state at M35. This modifies the topology of the E3 branch of the tree and the nomenclature of the corresponding hap- logroups, as shown in figure 1 (note that E3b now refers to all haplogroups with the M215 derived state). Five hundred fifteen haplogroup E3b subjects were iden- tified and further analyzed for the biallelic markers M34, M78, M81, M123, M281 (Underhill et al. 2000; Semino et al. 2002), and V6. The new V6 biallelic marker was discovered in the present survey in the TBL1Y gene by denaturing high-performance liquid chromatography analysis (primer sequences available on request). This marker identifies a subset of chromosomes previously assigned to E-M35* and now classified as E3b1e (fig. 1). No individual was found to carry the M281 muta- tion. We further typed 509 of the 515 E3b subjects for seven GATA STR (A7.1, A7.2, and A10 [White et al. 1999]; DYS19, DYS391, and DYS393 [Roewer et al. 1992, 1996]; and DYS439 [Ayub et al. 2000]) and four CA dinucleotide repeat (YCAIIa, YCAIIb, DYS413a, and DYS413b [Mathias et al. 1994; Malaspina et al. 1997]) polymorphisms. Both tetra- and dinucleotide mi- crosatellites were used to reconstruct haplogroup-spe- cific networks, through use of reduced-median and me.
dian-joining procedures (Bandelt et al. 1995, 1999). The seven tetranucleotide repeat polymorphisms were also used for the estimation of the time to the most recent common ancestor (TMRCA) (Goldstein et al. 1995; Slat- kin 1995; Thomas et al. 1998) and the time since two populations split from a common ancestor (TD estimator [Zhivotovsky et al. 2004]). For four of the tetranucle- otide loci here used, locus-specific mutation rates based on father-son transmissions (mi) are not available (Kayser et al. 2000). Since both TMRCA and TD estimations critically depend on the unknown parameter mi, we used the averaged effective mutation rate described by Zhi- votovsky et al. (2004), which is based on a list of markers close to the one used here. CIs for the TMRCA were obtained as described by Scozzari et al. (2001). It should be noted that uncertainties in the mutation rate, in the shape of the genealogy, and in the mutation process would increase the CIs. Since any two chromosomes sampled from two populations have a TMRCA older than the split between populations, and since we con- sidered as null the variance of the ancestral population at the time of its splitting, the figures reported here for the TD estimator represent upper bounds. In all of the analyses, except the networks, the YCAIIa, YCAIIb, DYS413a, and DYS413b dinucleotide repeats were not considered, since univocal assignment of phenotypic pat- terns to allelic series could not be obtained. We obtained an estimate of 25.6 thousand years (ky) (95% CI 24.327.4 ky) for the TMRCA of the 509 haplogroup E3b chromosomes, which is close to the 30 6 ky estimate for the age of the M35 mutation reported by Bosch et al. (2001) using a different method. Several observations point to eastern Africa as the home- land for haplogroup E3bthat is, it had (1) the highest number of different E3b clades (table 1), (2) a high fre- quency of this haplogroup and a high microsatellite di- versity, and, finally, (3) the exclusive presence of the un- differentiated E3b* paragroup. Our data show that haplogroup E3b appears as a collection of subclades with very different evolutionary histories. Haplogroup E-M78 was observed over a wide area, including eastern (21.5%) and northern (18.5%) Africa, the Near East (5.8%), and Europe (7.2%), where it represents by far the most common E3b subhaplo- group. The high frequency of this clade (table 1) and its high microsatellite diversity suggest that it originated in eastern Africa, 23.2 ky ago (95% CI 21.125.4 ky). The network of the E-M78 chromosomes reveals a strong geographic structuring, since each of the clusters a, b, and g (fig. 2B) reaches high frequencies in only one of the regions analyzed. Cluster a is largely characterized by the otherwise rare nine-repeat allele at A7.1 (we found only 3 such alleles out of 800 E[xE3b1] chro- mosomes analyzed [present study; R.S., unpublished data]), often associated with the uncommon DYS413
In conclusion, we detected the signatures of several distinct processes of migration and/or recurrent gene flow associated with the dispersal of haplogroup E3b lineages. Early events involved the dispersal of E-M78d
posted
A New Topology of the Human Y Chromosome Haplogroup E1b1 (E-P2) Revealed through the Use of Newly Characterized Binary Polymorphisms, 2011.
Beniamino Trombetta et al.
The new topology of the tree has important implications concerning the origin of haplogroup E1b1. Secondly, within E1b1b1 (E-M35), two haplogroups (E-V68 and E-V257) show similar phylogenetic and geographic structure, pointing to a genetic bridge between southern European and northern African Y chromosomes. Thirdly, most of the E1b1b1*(E-M35*) paragroup chromosomes are now marked by defining mutations, thus increasing the discriminative power of the haplogroup for use in human evolution and forensics.
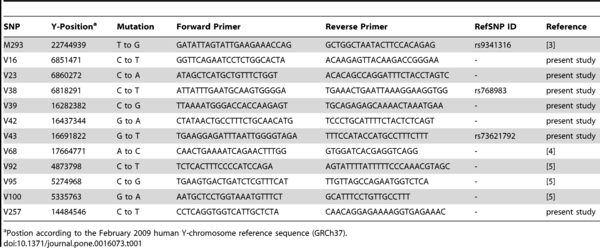
Within E-M35, there are striking parallels between two haplogroups, E-V68 and E-V257 [...] However, the absence of E-V68* and E-V257* in the Middle East (Table S2) makes a maritime spread between northern Africa and southern Europe a more plausible hypothesis.
Haplogroup E1b1 which is characterized by a high degree of internal diversity is the most represented Y chromosome haplogroup in Africa. Here we report on the characterization of 12 mutations within this haplogroup, eleven of which were discovered in the course of a resequencing and genotyping project performed in our laboratory. There are several changes compared to the most recently published Y chromosome tree [2]. Haplogroup E1b1 now contains two basal branches, E-V38 (E1b1a) and E-M215 (E1b1b), with V38/V100 joining the two previously separated lineages E-M2 (former E1b1a) and E-M329 (former E1b1c). Each of these two lineages has a peculiar geographic distribution. E-M2 is the most common haplogroup in sub-Saharan Africa, with frequency peaks in western (about 80%) and central Africa (about 60%).
Divorcing the Late Upper Palaeolithic demographic histories of mtDNA haplogroups M1 and U6 in Africa
Erwan Pennarun, Toomas Kivisild, Ene Metspalu, Mait Metspalu, Tuuli Reisberg, Doron M Behar, Sacha C Jones and Richard Villems
BMC Evolutionary Biology 2012, 12:234
"A Southwest Asian origin has been proposed for U6 and M1 [27-29]. Yet, this claim remains speculative unless some novel earlier Southwest Asian-specific clades, distinct from the known haplogroups, are found in which the described so far M1 and U6 lineages are nested. Claims for basal mutations shared with M1 have recently been made in the case of haplogroup M51 and M20 (both East Asian-specific clades [40,41]): They share a root mutation (C14110T) with M1.
However, one should be cautious with phylogenetic inferences drawn from these findings because this mutation is not unique in the phylogeny of mtDNA: it also occurs in the background of non-M haplogroups and therefore identity by descent within haplogroup M remains uncertain. Unfortunately, the sampling of extant populations of Africa and West Asia may not solve the question of their origin...
Assuming that M1 and U6 were introduced to Africa by a dispersal event from Asia, it would be difficult to accept their involvement in the first demographic spread of anatomically modern humans around 4045 KYA, as suggested by Olivieri et al. (2006), [29] who associated these two clades with the spread of Dabban industry in Africa. It has indeed been previously suggested that the colonisation of North Africa from the Levant took place during the early Upper Paleolithic, as marked by the Dabban industry in North Africa [42].
However, comparison of early Upper Palaeolithic artefacts from Haua Fteah and Ksar Akil does not support the notion that the early Dabban of Cyrenaica is an evidence of a population migration from the Levant into North Africa [43]. Marks [44] also noted differences between the two areas in terms of the methods of blade production, further arguing against a demographic connection between the regions. Likewise, the new coalescent date estimates for M1 obtained in this study are not compatible with the model implying the spread of M1 in Africa during the Early Upper Palaeolithic, 4045 KYA...
Our analyses do not support the model according to which mtDNA haplogroups M1 and U6 represent an early dispersal event of anatomically modern humans at around 4045 KYA in association with the spread of Dabban industry in North Africa as proposed earlier [28,29]. A West Asian origin for these haplogroups still remains a viable hypothesis as sister clades of U (and ancestral to it, macro-hg N (including R)) and M are spread overwhelmingly outside Africa, notably in Eurasia, even though the phylogeographic data on extant populations do not present a clear support for it.
No southwest Asian specific clades for M1 or U6 were discovered. U6 and M1 frequencies in North Africa, the Middle East and Europe do not follow similar patterns, and their sub-clade divisions do not appear to be compatible with their shared history reaching back to the Early Upper Palaeolithic." Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |






 UBBFriend: Email this page to someone!
UBBFriend: Email this page to someone!










 Printer-friendly view of this topic
Printer-friendly view of this topic