Indigenous Arabs are descendants of the earliest split from ancient Eurasian populations Juan L. Rodriguez-Flores 2016
(excerpts)
Abstract
An open question in the history of human migration is the identity of the earliest Eurasian populations that have left contemporary descendants. The Arabian Peninsula was the initial site of the out-of-Africa migrations that occurred between 125,000 and 60,000 yr ago, leading to the hypothesis that the first Eurasian populations were established on the Peninsula and that contemporary indigenous Arabs are direct descendants of these ancient peoples. To assess this hypothesis, we sequenced the entire genomes of 104 unrelated natives of the Arabian Peninsula at high coverage, including 56 of indigenous Arab ancestry. The indigenous Arab genomes defined a cluster distinct from other ancestral groups, and these genomes showed clear hallmarks of an ancient out-of-Africa bottleneck. Similar to other Middle Eastern populations, the indigenous Arabs had higher levels of Neanderthal admixture compared to Africans but had lower levels than Europeans and Asians. These levels of Neanderthal admixture are consistent with an early divergence of Arab ancestors after the out-of-Africa bottleneck but before the major Neanderthal admixture events in Europe and other regions of Eurasia. When compared to worldwide populations sampled in the 1000 Genomes Project, although the indigenous Arabs had a signal of admixture with Europeans, they clustered in a basal, outgroup position to all 1000 Genomes non-Africans when considering pairwise similarity across the entire genome. These results place indigenous Arabs as the most distant relatives of all other contemporary non-Africans and identify these people as direct descendants of the first Eurasian populations established by the out-of-Africa migrations.
All humans can trace their ancestry back to Africa [Cann et al. 1987], where the ancestors of anatomically modern humans first diverged from primates [Patterson et al. 2006], and then from archaic humans [Prüfer et al. 2014]. Humans began leaving Africa through a number of coastal routes, where estimates suggest these out-of-Africa migrations reached the Arabian Peninsula as early as 125,000 yr ago [Armitage et al. 2011] and as late as 60,000 yr ago [Henn et al. 2012]. After entering the Arabian Peninsula, human ancestors entered South Asia and spread to Australia [Rasmussen et al. 2011], Europe, and eventually, the Americas. The individuals in these migrations were the most direct ancestors of ancient non-African peoples, and they established the contemporary non-African populations recognized today [Cavalli-Sforza and Feldman 2003].
The relationship between contemporary Arab populations and these ancient human migrations is an open question [Lazaridis et al. 2014; Shriner et al. 2014]. Given that the Arabian Peninsula was an initial site of egress from Africa, one hypothesis is that the original out-of-Africa migrations established ancient populations on the peninsula that were direct ancestors of contemporary Arab populations [Lazaridis et al. 2014]. These people would therefore be direct descendants of the earliest split in the lineages that established Eurasian and other contemporary non-African populations [Armitage et al. 2011; Rasmussen et al. 2011; Henn et al. 2012; Lazaridis et al. 2014; Shriner et al. 2014]. If this hypothesis is correct, we would expect that there are contemporary, indigenous Arabs who are the most distant relatives of other Eurasians. To assess this hypothesis, we carried out deep-coverage genome sequencing of 104 unrelated natives of the Arabian Peninsula who are citizens of the nation of Qatar [Supplemental Fig. 1], including 56 of indigenous Bedouin ancestry who are the best representatives of autochthonous Arabs, and compared these genomes to contemporary genomes of Africa, Asia, Europe, and the Americas [The 1000 Genomes Project Consortium 2012; Lazaridis et al. 2014].
Figure 1. Principal component analysis [PCA] [Price et al. 2006] of the 104 Qatari genomes [circle], 1000 Genomes [triangle], and Human Origins [square] study samples. Shown are individuals plotted on principal components PC1 and PC2, with genomes color-coded by study and population, with the Q0 [Subpopulation Unassigned] in gray, Q1 [Bedouin] in red, Q2 [Persian-South Asian] in azure, and Q3 [African] in black. [A] Plot of all populations, defined by study and by population, in which all populations from the same region and study are grouped and color-coded together [1000 Genomes: Africa, America, East Asia, and Europe; Human Origins: Africa, America, Central Asia/Siberia, East Asia, Middle East, Oceania, South Asia, and West Eurasia]. [B] Plot of Middle Eastern subpopulations from Human Origins that cluster near Q1 [Bedouin] and Q2 [Persian-South Asian].
Figure 2. Y Chromosome [Chr Y] and mitochondrial DNA [mtDNA] haplogroup assignments. The Chr Y and mtDNA haplogroups were determined for Q1 [Bedouin], Q2 [Persian-South Asian], and Q3 [African]. [A] Pie charts of the haplogroup frequencies for Chr Y. [B] Pie charts of the haplogroup frequencies for mtDNA.
Admixture analysis The signal of an ancient bottleneck in the Q1 [Bedouin] is not unexpected given previous analyses of genomic admixture that found <1% African ancestry in this subpopulation [Omberg et al. 2012] and studies of worldwide population structure, which have inferred that the Q1 [Bedouin] genomes have the greatest proportion of Arab genetic ancestry, even when compared to Bedouins from outside Qatar and to Arabs in surrounding countries, including Yemen and Saudi Arabia [Hodgson et al. 2014; Shriner et al. 2014]. To confirm a similarly minute amount of African admixture for the Q1 [Bedouin] in our sample, we applied three methodologies: [1] an ADMIXTURE [Alexander et al. 2009] analysis of the genome-wide ancestry proportions in the 104 Qataris, the 1000 Genomes Project [The 1000 Genomes Project Consortium 2012], and Human Origins samples [Lazaridis et al. 2014]; [2] an ALDER [Loh et al. 2013] analysis of the proportion and timing of African ancestry in these same populations; and [3] a SupportMix [Omberg et al. 2012] analysis of the population assignments of local genomic segments of the 96 Q1 [Bedouin], Q2 [Persian-South Asian], or Q3 [African] Qatari genomes.
The ADMIXTURE analysis identified K = 12 ancestral populations as having the lowest cross-validation error [Supplemental Fig. 7A]. At this level of resolution, the Q1 [Bedouin] had a high average [84%] proportion of ancestry that was also present in the Human Origins Bedouin B population at a high average proportion [93%] [Supplemental Fig. 7B,C], in which this same ancestry was also shared with Saudis, and at lower levels among other Middle Eastern populations. This ancestry therefore appears to be the signal of an indigenous Arab ancestral population. The Bedouin A population also shared this ancestry but at a lower average proportion [45%] and appeared to be more admixed overall. The Q2 [Persian-South Asian] shared a large proportion [45% on average] of ancestry that dominates in Iranians [46% on average], consistent with a Persian ancestral population [Omberg et al. 2012]. The Q3 [African] shared the majority of ancestry with African populations as expected and were considerably admixed overall, again consistent with the known history of this subpopulation [Supplemental Fig. 7A; Omberg et al. 2012].
The ALDER analysis determined the relative percentage of African [Yoruba] ancestry in the Q1 [Bedouin] [2.6% ± 1.37] and Q2 [Persian-South Asian] [5.0% ± 1.41] at levels on par with estimates for other populations sampled in the region [Supplemental Fig. 8; Supplemental Table X], including Human Origins Bedouin and Saudi. This confirmed that recent African admixture is limited to the Q3 [African] subpopulation [37.6% ± 0.9], in which this estimate is on par with African American populations. An estimate of the timing of African admixture placed the number of generations for Q1 [Bedouin] [15.2] and Q2 [Persian-South Asian] [14.0] slightly higher than Q3 [African] [9.3], consistent with the Q1 [Bedouin] and Q2 [Persian-South Asian] reflecting more distant African admixture events and with the Q3 [African] reflecting the historical timing of the African slave trade in the region [Omberg et al. 2012].
The SupportMix analysis used six of the 1000 Genomes populations [two European, two Asian, and two African] [see Supplemental Methods for details] as ancestral proxy reference panels and produced a set of best guess admixture assignments based on highest similarity to these genomes. Although these 1000 Genomes populations do not include appropriate local populations most closely related to the Qataris needed for assessment of the true admixture composition of the genomes, the ancestry track length distribution of haplotypes assigned to African populations [Yoruba or Luhuya] provides a qualitative indicator of whether the subpopulations experienced recent admixture with African populations. As expected, the track lengths of the Q1 [Bedouin] and Q2 [Persian-South Asian] assigned to African 1000 Genomes populations were far shorter than those for Q3 [African] [Supplemental Fig. 9], again confirming that recent African admixture is limited to the Q3 [African] subpopulation.
Neanderthal ancestry We next analyzed Neanderthal admixture contributions to the ancestry of Q1 [Bedouin] compared to the Q2 [Persian-South Asian] and Q3 [African] Qataris, the 1000 Genomes populations, and the populations of the Human Origins samples using the F4 ratio and Patterson's D-statistic [Fig. 4; Supplemental Fig. 10, Supplemental Table XI; Patterson et al. 2012]. The results for both methods were highly correlated [Supplemental Fig. 10A]. The Q1 [Bedouin; F4 ratio = 0.026, D-statistic = 0.000] had more Neanderthal admixture than all African populations, including Q3 [African; F4 ratio range = −0.017 to 0.024, D-statistic range = −0.031 to −0.003]. The Q1 [Bedouin] also had Neanderthal admixture at levels comparable to Q2 [Persian-South Asian; F4 ratio = 0.024, D-statistic = −0.003] and to other Middle Eastern populations, including other Bedouin populations [Human Origins Bedouin A F4 ratio = 0.022, D-statistic = −0.003 and Bedouin B F4 ratio = 0.024, D-statistic = −0.003] and Saudi [F4 ratio = 0.026, D-statistic = −0.001]. Interestingly, the Q1 [Bedouin] did not tend to have higher Neanderthal admixture levels when considering populations outside of the Middle East, where the bulk of European populations had higher Neanderthal admixture [F4 ratio range = 0.018 to 0.041, D-statistic range = 0.003 to 0.010]. Yet, the percentage of Neandethal admixture with the Q1 [Bedouin] was higher than expected if it could be entirely explained by later admixture events between the Q1 [Bedouin] and Europeans [observed F4 ratio = 0.026 versus expected F4 ratio = 0.00247].
The higher Neanderthal ancestry in the Q1 [Bedouin] Qatari compared to African populations places the divergence of ancestral Arabs after the out-of-Africa bottleneck. Given the current evidence of the geographic range of Neanderthal populations stretching from Europe and the Mediterranean through Northern and Central Asia [Fu et al. 2014; Hershkovitz et al. 2015], the lower Neanderthal Ancestry in the Q1 [Bedouin] Qatari compared to populations within the ancestral Neanderthal range is also consistent with an early divergence of the ancestors of indigenous Arabs from other lineages that populated Asia and Europe. Yet, since the Neanderthal admixture in the Q1 [Bedouin] cannot be entirely explained by admixture with Europeans, this indicates there was some admixture between Neanderthals and ancestors of the Q1 [Bedouin] in the region of the Arabian Peninsula.
TreeMix analysis We also analyzed the autosomes of the combined 96 Q1 [Bedouin], Q2 [Perisan-South Asian] or Q3 [African] Qataris, and non-admixed populations of the 1000 Genomes Project using the population split and mixture inference method TreeMix [Pickrell and Pritchard 2012] to assess the relative genetic similarity of populations based on high-density, genome-wide allele frequencies. The analysis returned an overall tree for the 1000 Genomes populations that mirrored those found previously [Shriner et al. 2014] with the addition of the Q1 [Bedouin] and Q2 [Persian-South Asian] clustering on the branch that includes Europeans [Pérez-Miranda et al. 2006] and the Q3 [African] clustering with African populations [Fig. 5]. When migrations were allowed in the analysis, no migration events were observed between the Q1 [Bedouin] and African populations, even when allowing as many as five migration events [Supplemental Fig. 11]. These results are also consistent with what is known of the migration history of the Arabian Peninsula, including migration both to and from Europe during ancient and more recent eras of civilization, where this resulted in detectable admixture from European populations in both the Q1 [Bedouin] and Q2 [Persian-South Asian] [Omberg et al. 2012].
Figure 5. TreeMix [Pickrell and Pritchard 2012] hierarchical clustering analysis of the Q1 [Bedouin], Q2 [Persian-South Asian], and Q3 [African] and the 1000 Genomes Project samples. Shown is a maximum-likelihood tree of population splits inferred without subsequent migration events, in which branch lengths estimate divergence between populations [Europeans in shades of purple: CEU, FIN, GBR, IBS, TSI; East Asians in shades of brown: CHB, CHS, JPT; Africans in shades of orange: LWK, YRI, with the Q1 [Bedouin] in red, Q2 [Persian-South Asian] in azure, and Q3 [African] in black]. When allowing from one to five migration events in separate TreeMix analyses, none of the admixture loops connected the Q1 [Bedouin] with any African populations [Supplemental Fig. 10], consistent with the Q1 [Bedouin] having no recent African admixture.
Discussion
The hypothesis that the first Eurasian populations were established on the Arabian Peninsula and that contemporary indigenous Arabs are direct descendants of this ancient population is supported by two major conclusions derived from the combined evidence of this study. First, the analysis results for X/A diversity, the pairwise sequential Markov coalescent, genome-wide admixture, timing of African admixture, local admixture deconvolution, Neanderthal admixture, and application of TreeMix, support the inference that the Q1 [Bedouin] can trace the bulk of their ancestry back to the out-of-Africa migration events. Second, the combination of lower levels of Neanderthal admixture in the Q1 [Bedouin] than European/Asian populations and the outgroup position of the Q1 [Bedouin] compared to non-Africans in the pairwaise similarity clustering of high-density variants measured genome-wide, place the Q1 [Bedouin] as being the most distant relatives of other contemporary non-Africans. Given that the Q1 [Bedouin] have the greatest proportion of Arab genetic ancestry measured in contemporary populations [Hodgson et al. 2014; Shriner et al. 2014] and are among the best genetic representatives of the autochthonous population on the Arabian Peninsula, these two conclusions therefore point to the Bedouins being direct descendants of the earliest split after the out-of-Africa migration events that established a basal Eurasian population [Lazaridis et al. 2014]. This is also consistent with the majority of Q1 [Bedouin] being able to trace a significant portion of their autosomal ancestry through lineages that never left the peninsula after the out-of-Africa migration events since such deep ancestry would not be expected if the entire Arabian Peninsula population had been reestablished from Africa or a non-African population at a later point.
Given the complex history of migration patterns to and from European populations, and the complicated patterns of isolation and intra- and inter-marriage of the indigenous Bedouin populations [Hunter-Zinck et al. 2010; Sandridge et al. 2010], it is not surprising that among the Q1 [Bedouin] are individuals who retain an autosomal signal of being the most distant relatives of non-Africans, while population-level clustering based on migration-shifted allele frequencies places the Q1 [Bedouin] closer to Europeans. The basal position of the Q1 [Bedouin] also has interesting implications for theories about the frequency, timing, and path of major migration waves that established populations in Asia and Europe [Shi et al. 2008; Lazaridis et al. 2014; Shriner et al. 2014]. A few isolated Asian populations were previously suspected to be descendants of a separate out-of-Africa migration wave based on Y Chromosome data [Hammer et al. 1998; Shi et al. 2008]. Yet, distinct out-of-Africa migration events or separate migration waves emanating from the Arabian Peninsula into Europe and West Asia would be expected to place Bedouins/Europeans and Asians on separate branches of a pairwise clustering tree, distinct from our finding that places the Q1 [Bedouin] as direct descendants of the earliest lineage that split from the ancient non-African population.
A demographic scenario consistent with the evidence presented here is that the population ancestral to the Q1 [Bedouin] migrated out of Africa, and a subset of this population remained in the peninsula until the present day, while a second subset of this population migrated onward and colonized Eurasia. This migration scenario implies the signal of the same bottleneck would be present in all non-African populations, which has been observed thus far in coalescent analysis of contemporary non-African populations [Gronau et al. 2011; Fu et al. 2014; Schiffels and Durbin 2014] and for an anatomically modern human who lived 45,000 yr ago [Fu et al. 2014]. This is also consistent with the recent discovery of another anatomically modern human who lived 55,000 yr ago just northeast of the Arabian Peninsula that had morphological features similar to European peoples [Hershkovitz et al. 2015], where this individual could have been a descendant of the basal Eurasian population that remained on the peninsula. Under this migration scenario, although other waves of migration may have occurred, the descendants of these alternative waves either left no descendants or were integrated into the dominant populations.
Beyond the importance for disentangling human migration history, an early split of Eurasian lineages in the Arabian Peninsula has implications for the study of disease genetics for indigenous people in the region. For example, for a disease such as type 2 diabetes that has a prevalence of >18% in the Qatari population, associated genetic variants would not a priori be expected to be the same as those discovered in Europeans, when considering that indigenous Arabs are able to trace a significant portion of their ancestry back to ancient lineages on the Arabian Peninsula. More generally, this suggests that for any genome-wide association study [GWAS] or rare variant association study [RVAS] of diabetes or other complex diseases in Qatar, inference of deep ancestry in the Arabian Peninsula, using rare variation sampled by genome or exome sequencing, is critical for identifying new disease risk genes. Given the dearth of next generation sequencing studies conducted in Middle Eastern and Arab populations, these results indicate that a considerable number of variants that make important contributions to disease risk in these populations are yet to be discovered.
This study is the first analysis of Arabian Peninsula migration making use of deeply sequenced genomes from a sample of unrelated inhabitants of the peninsula. Although there have been many analyses of Chr Y and mtDNA sampled from Arab individuals [Abu-Amero et al. 2007, 2008, 2009; Rowold et al. 2007], and there have been previous surveys of genetic variation of people within the peninsula and immediately surrounding regions conducted with genotyping arrays [Behar et al. 2010; Hunter-Zinck et al. 2010; Alsmadi et al. 2013; Markus et al. 2014; Shriner et al. 2014] and deep exome sequencing [Rodriguez-Flores et al. 2012, 2014; Alsmadi et al. 2014], and by individual high-coverage genomes [Alsmadi et al. 2014; John et al. 2015], the sample of rare and common genetic variation throughout the genome in our sample provides a far more complete picture of how both ancient and recent migration events have contributed to the genetics of the modern peoples of the Arabian Peninsula. For understanding how human migration history has determined the structure of modern genomes, our identification of a cluster of Q1 [Bedouin] as the most distant ancestors of non-Africans is of considerable interest, particularly given the suspected route of migration out of Africa and into the surrounding continents. The possibility that the Q1 [Bedouin] are descendants of the first Eurasians provides an additional piece of the puzzle concerning ancient migration routes and the establishment of ancient non-African populations.
Inclusion criteria Qatar is a peninsula nation on the eastern edge of the Arabian Peninsula [Supplemental Fig. 1]. The population of Qatar includes more than 2 million inhabitants, comprised of ∼300,000 nationals with roots in Qatar predating the discovery of oil and gas and establishment of an independent nation in 1970 and the more than 1.7 million immigrants who mostly arrived in the past decade [Qatar Statistics Authority 2013, http://www.qsa.gov.qa/QatarCensus/Pdf/Population above 15 by educational attainment, nationality, age, sex and marital status.pdf]. As selection criteria, we required that subjects be third-generation Qataris and all ancestors were Qatari citizens born in Qatar, as assessed by questionnaires. Recent immigrants or residents of Qatar who traced their recent ancestry to other geographic regions were excluded.
Natives of the Arabian Peninsula can be divided into at least three genetic subpopulations that reflect the historical migration patterns in the region: Q1 [Bedouin], Q2 [Persian-South Asian], and Q3 [African] [Hunter-Zinck et al. 2010; Omberg et al. 2012; Rodriguez-Flores et al. 2012]. A panel of 48 SNPs was genotyped by TaqMan [Life Technologies] sufficient for classification into one of the three subpopulations based on >70% ancestry in one cluster in a STRUCTURE analysis with k = 3 used to identify individuals that could unambiguously be placed in one of these three groups [Supplemental Fig. 2; Pritchard et al. 2000; Rodriguez-Flores et al. 2012]. Our primary focus was the Q1 [Bedouin] genetic subpopulation because of its deepest ancestry in Arabia [Ferdinand et al. 1993], so we selected 60 Q1 [Bedouin] individuals to include in the sample. We additionally selected 20 Q2 [Persian-South Asian] and 20 Q3 [African] to use as controls in the analysis, and an additional eight Q0 [Subpopulation Unassigned] individuals that could not be confidently placed in one of these subpopulations, defined as not having >70% ancestry in any of the three groups as determined by STRUCTURE. The total sample therefore included 108 individuals with an even distribution of males and females [see Supplemental Methods; Supplemental Table I].
Illumina deep sequencing of the genomes In order to characterize the spectrum of genetic variation, each of the 108 Qatari genomes were sequenced to a median depth of 37× [minimum 30×] through the Illumina Genome Network [see Supplemental Methods for details].
Relatedness among Qataris Given the high rate of consanguineous marriage previously reported in the Qatari population [Hunter-Zinck et al. 2010; Mezzavilla et al. 2015], we sought to quantify the relatedness between individuals in our sample and to exclude closely related individuals that could potentially confound population genetics analysis methods that assume the input sample is unrelated. In order to conduct the relatedness analysis, autosomal SNPs in 108 Qatari genomes [described above] were filtered using PLINK 1.9 [Chang et al. 2015], and relatedness between the 108 Qatari genomes was assessed using kinship coefficients estimated by KING-robust [Manichaikul et al. 2010] and PREST-plus [McPeek and Sun 2000] [see Supplemental Methods]. Both methods found the same five first-degree and second-degree relationships, in which these relationships were then confirmed by investigative reassessment of medical records. One individual from each of the five pairs of relatives was then excluded from the study. Three of the pairs of relatives formed a trio; hence, two individuals were excluded from the trio, and one individual was excluded from each of the other two pairs, resulting in exclusion of four relatives in total.
Integration with the 1000 Genomes Project Phase 1 An integrated SNP call set was produced for ancestry analysis for a total of 1200 genomes, combining the 108 Qatari genomes with the 1092 genomes from the 1000 Genomes Project Phase 1 [1000 Genomes] [The 1000 Genomes Project Consortium 2012] [see Supplemental Methods]. The integrated call set included 11,711,411 autosomal biallelic SNPs. The transition:transversion ratio of this final set was 2.2, close to values previously observed in the 1000 Genomes Project [The 1000 Genomes Project Consortium 2012]. Based on the concordance and quality measures, the calls generated from our pipeline were considered to be high quality, and these were used for all further aspects of this study. After exclusion of four related Qataris [Supplemental Table III], the final integrated call set included 11,711,386 autosomal biallelic SNPs in 1196 genomes.
Integration with Human Origins data set The 1000 Genomes Project Phase 1 is an excellent resource for rare variant discovery; however, it is limited in terms of the breadth of global populations sampled. Unfortunately, at the time of writing, no global resource of sequenced genomes existed; hence, the next best alternative for comparison of the Qataris to populations around the world is the Human Origins Fully Public Dataset [referred to here as Human Origins [HO]], which includes genotype data for 1917 indivduals from Africa, West Eurasia [including Middle East], South Asia, East Asia, Central Asia/Siberia, and America. In particular, the West Eurasian, African, and South Asian data sets include populations sampled in countries close to Qatar, where detection of shared ancestry is of interest in this study. The data set also includes data from archaic genomes, such as Altai Neanderthal, Denisova, and chimpanzee, which are of interest in this study for quantification of Neanderthal ancestry. The Human Origins data set includes a number of samples also present in the 1000 Genomes Project [Supplemental Table IV], and for these samples, the Human Origins overlap data is kept.
In order to conduct population genetic analysis on a combined data set of the 104 Qatari genomes [QG, n = 104], the 1000 Genomes Project Phase 1 [1000G-HO, n = 1028 after exclusion of duplicates], and Human Origins Fully Public Dataset [HO, n = 1862 after exclusion of archaic genomes, ancient genomes, and other genomes not relevant to this study] [Supplemental Table V], a set of sites overlapping between the integrated Qatari genomes plus the 1000 Genomes Project minus Human Origins, and the Human Origins data set were identified. Of 600,841 SNPs in the Human Origins data set and 11,711,386 SNPs in the Qatari genomes plus the 1000 Genomes Project data set, 388,805 SNPs overlapped. Further filtering was conducted on the data set, pruning SNPs based on linkage disequilibrium using PLINK [Purcell et al. 2007], --indep-pairwise 200 25 0.4, matching parameters used previously [Lazaridis et al. 2014]. After linkage disequilibrium-pruning, the final data set for analysis included 197,714 SNPs segregating in the three data sets [QG, 1000G-HO, and HO].
Inbreeding coefficient In order to place the high reported consanguinity in Qatar in a global context, the inbreeding coefficient was calculated using PLINK 1.9 [Chang et al. 2015] for Q1 [Bedouin], Q2 [Persian-South Asian], and Q3 [African] Qataris, the 1000 Genomes Project minus Human Origins overlap, and Human Origins populations [see Supplemental Methods].
Principal component analysis A PCA [Price et al. 2006] was carried out for the combined 104 Qatari genomes, the 1000 Genomes Project minus Human Origins overlap, and Human Origins samples using the 197,714 SNPs in the integrated data set [filtering criteria described above]. Using the results of this large-scale analysis, visual assessment of clustering and population overlap was used to confirm expected relationships between the analyzed populations. Four distinct plots of a single PCA run were constructed: one comparing the Qatari genomes to the 1000 Genomes populations [Supplemental Fig. 5A], one comparing Qataris to the 1000 Genomes and Human Origins Samples including two visualizations of the full data set [Fig. 1A, color-coded by regional meta-populations; Supplemental Fig. 5B, color-coded by detailed population], and one comparing Qataris to Middle Eastern populations from the Human Origins data set [Fig. 1B]. For the latter, in order to compare Qataris to Middle Eastern populations with potential for recent shared Bedouin ancestry with Qataris sampled by the Human Origins data set, populations from the Middle East previously labeled in Lazaridis et al. [2014] as West Eurasia, were relabeled as Middle East, including Bedouin A, Bedouin B, Druze, Egyptian Comas, Egyptian Metspalu, Iranian, Jordanian, Lebanese, Palestinian, Saudi, Syrian, Turkish, Turkish Adana, Turkish Aydin, Turkish Balikesir, Turkish Istanbul, Turkish Kayseri, Turkish Trabzon, and Yemen.
Y and mitochondria haplogroup assignment In order to determine the prevalence of known Chr Y and mtDNA haplogroups in Qatar, SNP genotypes were generated simultaneously for the 108 Qatari genomes using an updated version of GATK [v3.1.1] [DePristo et al. 2011] that supports haploid chromosome calling [n = 53 Chr Y, n = 108 mtDNA]. For one of the genomes, the sample was originally thought to be male but is most likely female due to low call rates on Chr Y. This sample was excluded from Chr Y analysis and X/A diversity analysis, but was included in autosomal and mtDNA analysis. Mean coverage of mapped reads was 11× in Chr Y and 3892× in mtDNA. After exclusion of related and Q0 [Subpopulation Unassigned] [admixed] Qataris, the remaining samples included 47 Chr Y and 96 mtDNA.
Haplogroup assignments for the Chr Y and mtDNA were made using previously characterized variants. For Chr Y, these assignments were made using YFitter [Jostins et al. 2014] by using variants limited to known SNPs cataloged by the International Society of Genetic Genealogy [Jobling and Tyler-Smith 2003] within a 10-Mb interval of the Y Chromosome that is known to be amenable to analysis based on short read sequencing [Skaletsky et al. 2003; Poznik et al. 2013]. For mtDNA, these assignments were made using HaploGrep [Kloss-Brandstätter et al. 2011] by using the set of known haplogroup-specific variants in the PhyloTree [van Oven and Kayser 2009] database.
In order to quantify the differences between mtDNA and Chr Y in terms of diversity of the haplogroups identified, the proportion of variance among and within populations was quantified for Chr Y and mtDNA using the AMOVA function in Arlequin [Supplemental Methods; Excoffier et al. 1992; Excoffier and Lischer 2010]. The analysis was repeated eight times, including separate analysis of Chr Y and mtDNA, for three-way comparison of the populations, as well as all possible two-way comparisons [Q1/Q2, Q1/Q3, Q2/Q3]. The proportion of variance among and within populations was tabulated, as well as the estimated Fst and P-value for both.
Comparison of X Chromosome to autosomal diversity The ratio of X-linked to autosomal nucleotide diversity [X/A] for different populations was computed following the approach in Gottipati et al. [2011] and Arbiza et al. [2014] [Supplemental Methods].
Coalescent analysis To infer the extent and timing of bottlenecks, the pairwise sequential Markov coalescent [PSMC] [Li and Durbin 2011] was applied to the 96 Q1 [Bedouin], Q2 [Persian-South Asian], or Q3 [African] Qatari genomes. A plot of effective population size versus years in the past was generated for each of the genome using instructions from the PSMC manual [Li and Durbin 2011; see Supplemental Methods]. For comparison, the same PSMC pipeline was run on BAM files of Illumina deep sequencing reads mapped to the GRCh37 human reference genome for an individual of European ancestry [NA12878, Utah resident with Northern and Western European ancestry, CEU] and an individual of African ancestry [NA19239, Yoruba in Ibadan, Nigeria, YRI] sequenced as part of the 1000 Genomes Pilot [The 1000 Genomes Project Consortium 2010]. The resulting PSMC plots for these two individuals were shifted slightly, such that they align with Qatari PSMC plots at distant [>200,000 yr ago] timescales [Fu et al. 2014].
Genome-wide admixture analysis In order to learn more about the ancestry of the sampled Qataris, a genome-wide admixture analysis was conducted on the combined data set of 104 Qatari genomes, the 1000 Genomes Project minus Human Origins overlap, and Human Origins using ADMIXTURE [Supplemental Methods; Alexander et al. 2009]. The cross-validation error was calculated for a range of expected number of ancestral populations [K], and the K with the lowest cross-validation error was used to quantify ancestry, in this case K = 12.
African admixture proportion and timing In order to estimate the proportion and timing of African admixture in Qatari populations, the genomes of Qataris and world populations were analyzed using ALDER 1.2 [Supplemental Methods; Loh et al. 2013].
Local admixture analysis An admixture deconvolution analysis was performed on the 96 Q1 [Bedouin], Q2 [Persian-South Asian], or Q3 [African] Qatari genomes using the 11,711,386 autosomal SNPs segregating in both the 1000 Genomes Project and Qatari genomes using SupportMix [Supplemental Fig. 9; Supplemental Methods; Omberg et al. 2012].
Neanderthal ancestry In order to compare the proportion of Neanderthal admixture in Q1 [Bedouin] Qataris with that of other populations in the 1000 Genomes Project [The 1000 Genomes Project Consortium 2012] and Human Origins [Lazaridis et al. 2014], the F4 ratio [Patterson et al. 2012] and Patterson's D-statistic [Patterson et al. 2012] were estimated using the qpF4ratio and qpDstat programs, respectively, from the ADMIXTOOLS 3.0 package [Supplemental Methods; Patterson et al. 2012].
We additionally considered the expected F4 ratio for the Q1 [Bedouin] under the scenario of no admixture between Neanderthal and direct ancestors of Q1 [Bedouin], such that observed Neanderthal ancestry in Q1 [Bedouin] would be entirely due to European admixture. From the estimated components of the ADMIXTURE analysis with K = 12, the Southern European ancestry in the Q1 [Bedouin] is 8.2% on average, and the Northern European ancestry in Q1 [Bedouin] is 1.3% on average, totaling 9.5% of the genome. If the Q1 [Bedouin] had never mixed with Neanderthal prior to introduction of European admixture, assuming no selection against introgressed genomic intervals, we would therefore expect an F4 ratio in Q1 [Bedouin] to be on the order of 1/10 of those observed in European populations.
TreeMix analysis We performed a TreeMix analysis [Pickrell and Pritchard 2012] of the 96 Q1 [Bedouin], Q2 [Persian-South Asian], or Q3 [African] Qatari genomes and the 1000 Genomes Project excluding admixed populations [Puerto Rican, Mexican, Colombian, and African Ancestry in Southwest US] [Supplemental Methods].
Neighbor-joining tree clustering In order to determine if any of the Qatari genomes were the most distant ancestors of all non-African populations, neighbor-joining trees were constructed for the 104 Qatari genomes and the 1000 Genomes Project using the 11,711,386 autosomal SNPs segregating in both data sets. For each pair of genomes, the proportion of shared alleles [PSA] [Mountain and Cavalli-Sforza 1997], or 1 minus the proportion of the genome identical by state [IBS], was calculated using the --distance -square -1-ibs function in PLINK 1.9 [Purcell et al. 2007; Chang et al. 2015], which outputs a 1196×1196 matrix of distances [1 minus IBS distance or PSA]. A neighbor-joining [NJ] tree was constructed using a recently updated version of the original NJ [Saitou and Nei 1987] algorithm called NJS [Criscuolo and Gascuel 2008] that is better at handling missing values, as implemented in the APE package in R [Paradis et al. 2004; R Core Team 2014]. Overall, this approach is computationally tractable for millions of markers genotyped in thousands of genomes and produces similar topologies to maximum-likelihood clustering methods but requires only a fraction of the compute time, where the trade-off is a sacrifice in the accuracy of branch lengths [Tateno et al. 1994]. The algorithm takes the distance matrix as input and outputs a tree. In order to confirm the robustness to sample ordering, the order of samples in the matrix was shuffled and reclustered 100 times, in which all reclusterings recovered the same tree. In order to produce bootstrap support values for the tree, 100 reclusterings of the tree were generated based on random sampling of SNPs. For each bootstrap iteration, 11,711,386 random [with replacement] SNPs were selected using a Python script [www.python.org], and then the PSA distance matrix and NJ tree were recalculated using these SNPs. Bootstrap support was calculated using the Python package SumTrees [Sukumaran and Holder 2010].
For visualization, the tree was rooted at the most recent common ancestor [MRCA] node of the largest cluster of the 1000 Genomes Yoruba [YRI] genomes in the tree. A color version of the tree was produced using TreeGraph 2 [Stöver and Müller 2010] by manually coloring the branches leading to each node. A single color is assigned to each population, with populations from the same continent having similar colors: Europeans in shades of purple, Asians in shades of brown, Americans in shades of green, Africans in shades of orange, Q1 [Bedouin] in red, Q2 [Persian-South Asian] in blue, Q3 [Sub-Saharan African] in black, and Q0 [Subpopulation Unassigned] in gray. When a cluster of nodes includes different populations, the terminal branches were given population-specific colors, whereas the shared higher-order branches for the cluster were given the color of the population in majority. For example, if 10 Q1 [Bedouin] and 1 Q0 [Subpopulation Unassigned] were in a cluster, the branches above where the nodes come together were colored red.
Posts: 42918 | From: , | Registered: Jan 2010
| IP: Logged |
posted
Who bunch of nobodies. No one well known penned their name to this study.
When recent paper 2016. Similar paper have been written which contradict their hypothesis posted on ESR. They used YRI but not North Africans. Everyone knows Bedoiuns are North Africans.
What premise/ojective is to identify who the Bediouns are in relation to Africans and Europeans. They are after basal Eurasian and the Neolithics. I told you this is where the new war is at. They have ceded AEians to Africans now they bendng over backwards to wrestle Neolithics from Africa. They dont want to be classified as subset of Africans. Ooops! Too late. Lol!
Where oddly they chose Qatar to sample. Same population and dataset as Henn infamous back-migration paper. As I told many people here the Bedoiuns ar ethe indigenous popualtion of the Arabian Peninsular NOT Turks of the Levant. -----
I notice they use TreeMix but they have arrows going FROM Q1 to Africa. Anyone can explain why?
Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
quote:Originally posted by xyyman: now they bendng over backs to wrestle Neolithics from Africa. They dont want to be classified as subset of Africans. Ooops! Too late. Lol!
Give your definition of neolithic. Then name the culture or cultures that are neolithic and time period.
Show us you're not bluffing
Posts: 42918 | From: , | Registered: Jan 2010
| IP: Logged |
posted
I double dare you. lol!.....really. anyways.
==
-------- Quote: The higher Neanderthal ancestry in the Q1 (Bedouin) Qatari compared to African populations places the divergence of ancestral Arabs ***after*** the out-of-Africa bottleneck.
When migrations were allowed in the analysis, no migration events were observed between the Q1 (Bedouin) and African populations, even when allowing as many as five migration events (Supplemental Fig. 11).
The Arab haplogroup J1 was the dominant haplogroup in the Q1 (Bedouin) Qataris, but this haplogroup was not represented at all among the Q2 (Persian-South Asian) Qataris (Fig. 2A). This confirmed that these are genetically well defined subpopulations that are relatively isolated from one another (Omberg et al. 2012).
We also confirmed these clusterings just with the 104 Qataris and 1000 Genomes samples based on the same set of autosomal SNPs (Supplemental Fig. 5B). These analyses reproduced the population clustering observed previously (Hunter-Zinck et al. 2010; Omberg et al. 2012), with the Q1 (Bedouin) closest to Europeans, the Q2 (Persian-South Asian) between Q1 (Bedouin) and Asians, and the Q3 (African) closest
to African populations. A plot of just the Middle Eastern populations on the principal components also showed clustering as expected, with the Q1 (Bedouin) clustering with previously sampled Bedouins and Arabs, Q2 (Persian-South Asians) with Iranians, and Q3 (African) outside of the Middle Eastern cluster ***(data not shown)**** (Fig. 1B). ---------- Xxyman Comments - Up to their old tricks again Like Busby. DATA NOT SHOWN. Also interesting is that Bedoiun (Q1) is closest to Europeans but do NOT carry primarily J1. Really lol! BTW they also carry R-V88. You guessed it. Sibling to R1b! North Africans carry primarily J1. Now why would there be no migration between Bediouns and YRI when Lazaridis was explicit when he stated he could NOT disentangle YRI and Bedoiuns. I know why, because they are one and the same. Tada!
-------------------- Without data you are just another person with an opinion - Deming Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
quote:Originally posted by xyyman: now they bendng over backs to wrestle Neolithics from Africa. They dont want to be classified as subset of Africans. Ooops! Too late. Lol!
Give your definition of neolithic. Then name the culture or cultures that are neolithic and time period.
Show us you're not bluffing
^no answer from xyyman As I suspected. xyyman doesn't know what neolithic means and he has no examples of neolithic sites. He just likes the way the word sounds
Posts: 42918 | From: , | Registered: Jan 2010
| IP: Logged |
I'm Skeptical "Indigenous Arabs" Are Basal Eurasians
This is a good paper. Theyve taken a stab at it, and are very circumspect. But in the end they state that these two conclusions therefore point to the Bedouins being direct descendants of the earliest split after the out-of-Africa migration events that established a basal Eurasian population.
To catch everyone up, Lazaridis et al. suggested based on results from ancient DNA that many West Eurasian populations have an ancestry which derives from a lineage basal to all other non-Africans unmixed with this population. That means that the genetic distance of this group to Pleistocene European hunter-gatherers and Pleistocene Australians is the same, while the genetic distance between these two groups is smaller than between them and this population. Therefore they are termed basal Eurasians, or bEu.
But it is also important to note that they are a construct. The ancient DNA has not found any unmixed basal Eurasians. This is in contrast to other groups which are used as donor populations: European hunter-gatherers and Siberian hunter-gatherers. About ~50% or so of the ancestry of the Anatolian farmers who were the precursor of the first agriculturalists in Europe derive from bEu ancestry, with the balance consisting of a heritage similar to to European hunter-gatherers. The hunter-gatherers recently discovered in the Caucasus also have this bEu ancestry. Ergo, almost all West Eurasian and South Asian populations have bEu ancestry.
In the paper above, which is open access, the authors found a group of Qatari Bedouin, who seem to have low admixture from Africans or other Middle Eastern groups. Though some preliminary analysis was done with SNP-chips, they went whole genome for most of the work (allowing them to look for rare variants, etc.). I would have been convinced to a great extent if they put a TreeMix graph out which showed that their indigenous Arab population was a good donor to ancient Anatolians along with European hunter-gatherers. But I did not see that. Or they could have done an F4 ratio test showing that the Bedouin were more basal Eurasian than any other modern population. I did not see that.
I did see an F4 ratio test for Neanderthal admixture. I am not confident that their assertions hold. Take a look at the pattern of Neanderthal admixture in the supplements; its all over the place. It isnt in line with the broad patterns found in the latest work out of David Reichs lab.
There are also some assumptions within the paper which I think are untenable. They seem to be positing a continuity of these Qatari Bedouin within the Arabian peninsula for tens of thousands of years. The divergence of the bEu population, putatively ancestral to these Bedouin, occurred from other non-Africans even before the settlement of Australia, over 50,000 years ago! I dont think it is likely that the Bedouin were resident in or around the Arabian peninsula for that long.
Finally, theres some reference to effective population sizes vs. X and autosome. This isnt a major part of the paper, but I would be skeptical of these sorts of claims. There is a lot of work in this area, and it turns out everything is way more clouded than you might think on first blush.
Overall, good paper. But theres still a mystery here. The only solution is clearly more ancient DNA from this region.
Posts: 42918 | From: , | Registered: Jan 2010
| IP: Logged |
posted
I believe the contemporary Bedoiuns are not the original Arabians they are a mixture of white groups including the Turks, Gutians and People of the Sea.
-------------------- C. A. Winters Posts: 13012 | From: Chicago | Registered: Jan 2006
| IP: Logged |
Some say that the Mahra people (Arab Salah tribe) (Mehri language speakers) might have a deeper history than others in the region
however the Arabian peninsula has undergone many aridization periods where people may have been migrating in and out
Posts: 42918 | From: , | Registered: Jan 2010
| IP: Logged |
quoteQuote: The higher Neanderthal ancestry in the Q1 (Bedouin) Qatari compared to African populations places the divergence of ancestral Arabs ***after*** the out-of-Africa bottleneck.
===
why? Just like early farmers (Lazaridis et al 2016). Bedouins carry very little more neanderthal ancestry than extant Africans. So, yes, Bediouns are a primarily Africans who admixed with first OOA. That would explain the slight increase in "Neanderthal" ancestry compared to more distant population like Asians...and Northern Europeans.
This is not rocket science.
-------------------- Without data you are just another person with an opinion - Deming Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
posted
And if I were a betting man I would guess Bedouins and indigenous North Africans carry the same frequency of Neanderthal ancestry.
-------------------- Without data you are just another person with an opinion - Deming Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
Rodriguez-Flores et al 2016 in their study about the ancient Eurasian population split sequenced with a median depth of 37× 108 purportedly unrelated individuals who are citizens of the nation of Qatar. 56 of them are of indigenous Bedouin ancestry who are the best representatives of autochthonous Arabs. Genetically most of them could be placed in three distinct clusters: 60 Q1 (Bedouin), 20 Q2 (Persian-South Asian), and 20 Q3 (African), as well as 8 Q0 (Subpopulation Unassigned). 53 of those have Qataris have Y chromosomes and the Raw data (SRA) is available. The study analysis shows that J2 is present in all three population clusters with a major presence (ca. 1/3) in the Persian-South Asian group. I hope YFull is going to analyze those and include them in their YTree.
According to the current grouping in J2
@FTDNA project we have those lineages in Qatar: J2a-M47 (1) J2a-M47>Y14678 (2) J2a-M67>? (3) J2a-M67>Z500>M92 (2) J2a-M319 (1) J2a-L24>L25>Z7700 (4) J2a-L24>L25>? (2) J2a-PF5197>PF5172>PF5191>PF5174>S19231 (3) J2a-Z6064>Z6063>Z6057>SK1366>M68 (1) J2a-Z6064>PH4959 (1) J2b1-M205>PH4306 (1) J2b2-Z1827>Z2456 (1) _______________________________________________________
American Journal of Cardiology, compared the genomes of Qatars sub-Saharan African subgroup with the Persian-South Asian group and the Bedouin by analyzing 228 individuals. The investigators found that 17 percent of the African-derived subgroup had the R145C variant, while none of the Bedouin or Persian participants had the mutation. The research team then examined the 1000G data and found that the R145C variant is virtually non-existent among populations of non-African descent but is found in between five and 12 percent of African-derived populations. A study of 1,266 African-Americans in the New York area found that four percent carried the R145C variant.
__________________
Prevalence of the Apolipoprotein E Arg145Cys Dyslipidemia At-Risk Polymorphism in African-Derived Populations Jan 2014
Maen D. Abou Ziki, BA, Yael Strulovici-Barel, MS, Neil R. Hackett, PhD, Juan L. Rodriguez-Flores, PhD, Jason G. Mezey, PhD, Jacqueline Salit, MS, Sharon Radisch, MS, Charleen Hollmann, PhD, Lotfi Chouchane, MD, Joel Malek, PhD, Mahmoud A. Zirie, MD, Amin Jayyuosi, MD, Antonio M. Gotto Jr., MD, DPhil, Ronald G. Crystal, MD
Posts: 42918 | From: , | Registered: Jan 2010
| IP: Logged |
quote:Originally posted by xyyman: now they bendng over backs to wrestle Neolithics from Africa. They dont want to be classified as subset of Africans. Ooops! Too late. Lol!
Give your definition of neolithic. Then name the culture or cultures that are neolithic and time period.
Show us you're not bluffing
^no answer from xyyman As I suspected. xyyman doesn't know what neolithic means and he has no examples of neolithic sites. He just likes the way the word sounds
What is the Q assignment?
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
"The Turkic languages originated in a region spanning Western China to Mongolia from where it expanded to Central Asia and farther west.[3]"
Isn't Hebrew and Arabic AfroAsiatic languages?
quote:Originally posted by DD'eDeN: Xyyman: "As I told many people here the Bedoiuns ar ethe indigenous popualtion of the Arabian Peninsular NOT Turks of the Levant."
Hmm. Linguistically, Hebrew & Arabic are basally tied to Turkic.
Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
posted
From: Kuwaiti population subgroup of nomadic Bedouin ancestryWhole genome sequence and analysis Sumi Elsa John 1,
quote "273 Kuwaiti natives, we recently demonstrated three distinct genetic subgroups in Kuwaiti population [2]: Kuwait P (KWP) of Persian ancestry; Kuwait S (KWS) of city-dwelling Saudi Arabian tribe ancestry, and Kuwait B (KWB) that includes most of the tent-dwelling Bedouin participants (recruited to provide samples for genotyping). The KWB is distinguished from the other two groups by a characteristic presence of 17% African ancestry (ranging from 11.7% to ***39.4%**)"
---- By African they mean SSA not including North Africans. Include North Africans and ...well...you know the rest
-------------------- Without data you are just another person with an opinion - Deming Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
posted
So as anyone can see. Kuwait, Qatar, Levant, Yemen etc. All the Bedouins are predominantly Africans....no slavery needed. lol!
Quote: Arabian ancestry is seen more in the Saudi Arabian tribe ancestry subgroup (at 69%) than in the Bedouin group (at 40%). Populations from other states of the Arabian Peninsula also display such a ***characteristic presence*** of African ancestry:
------------- Of course by "Arabian" vs African they mean North African cf to SSA.
THE TURKS WERE NOT INCLUDED!!!!!! The dominant Turks of the Levant and Arabia are not indigenous!!! FYI The Turks are those many visualize as "Arabs". they are not Arabs.
Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
posted
So Sudan has the highest frequency of J1e in the region. Significance? ---------- Quote: "The KWB sample is observed to have J1e [J-P58] Y-chromosome haplogroup which is seen in the Arabian Peninsula. The overall estimated time of expansion of J1e haplogroup is around 10,000 years and the ancestors of J1e haplogroups are observed in the Caucasus and eastern Anatolian populations [12]. The frequencies of J1e in populations from the region are [12]: Sudan: (74.2%; n = 35); Yemen (67.7%; n = 62); Negev Bedouin (64.4%; n = 28); Ismaili Damascus (58.8%; n = 51); Qatar (56.9%; n = 72); Jordan (48.7%; n = 76); Sunni Hama (44.4%; n = 36); Oman (37.2%; n = 121); and UAE (34.8%; n = 164). The observed high value of 67.7% for the frequency of J1e in Yemeni population"
" Examination of genetic clusters derived using principal component analysis (PCA) for Kuwait population (Fig. 1) reveals that this sample is located deep in the Bedouin cluster (and not at boundaries of the clusters or in regions that overlap among the three clusters)."
" The mitochondrial haplogroup (indication of maternal ancestry) of the Bedouin participant is determined as L3d1a1a [L3d], that is predominantly seen in West-Central Africaamong the Fulani [13], Chadians [13], Ethiopians [14], Akan people [15], Mozambique [14], and Yemen [14]."
-------------------- Without data you are just another person with an opinion - Deming Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
posted
Obviously the geographic pattern is clear. The Sahel of Africa comprising Africans peoples starts at the coast of West Africa and ends at the West Coast of Persia/Iran....and even beyond. Incldunding all of the Arabian Peninsular. While North Africa extends into Sardinia, Crete, Malta, Sicily, Corsica and the tips of main land Europe. That is why southern Europeans have so much African Ancestry. These new Neo(new) Africans migrated and admixed with the first OOA metapopulation heavily relate dto the Onge, Andamans and other Black Asians.
-------------------- Without data you are just another person with an opinion - Deming Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
I know what Arabic sounds like, I know what Hebrew sounds like and I know what Turkish sounds like. But never heard of this.
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
posted
I'll have to find the table, from Merritt Ruhlen's book The Origin of Language.
You (and most others) will probably dispute that the Turkic words are too different from the Arabic and Hebrew.
However, it is the consistency that is important, since through time all words evolve (sound or meaning) away from their root (an analogy: tree grows away from seed, to prevent competing with the original).
[I am aware that Turkic is Altaic; Semitic & Arabic are Afro-Asiatic.]
quote:Originally posted by DD'eDeN: I'll have to find the table, from Merritt Ruhlen's book The Origin of Language.
You (and most others) will probably dispute that the Turkic words are too different from the Arabic and Hebrew.
However, it is the consistency that is important, since through time all words evolve (sound or meaning) away from their root (an analogy: tree grows away from seed, to prevent competing with the original).
[I am aware that Turkic is Altaic; Semitic & Arabic are Afro-Asiatic.]
Turkic perhaps finds influence of Arabic due to the spread of Islam. Other than that I see and hear no relation. Turkish people do have words which sound similar to Iranian Farsi, which is rooted in PIE. I heard friends talk about this. Turkish and Iranian. They didn't know about linguistics and substratums. But I did explain it to them. The conversation was about meat, and several words they have for meat. Which in some parts (communities) and dialects had a very basic meaning and understanding.
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
posted
Yes, the eastward spread of Islam (and Arabic), and the westward spread of Mongols power and southward spread of Mughals (speaking Turkic but adopting Farsi(Avestani) in hierarchical ceremonial/legal matters), lots of mixture.
posted
[]The KWB is distinguished from the other two groups by a ****CHARACTERISTIC**** presence of 17% African ancestry (ranging from 11.7% to ***39.4%**)"
quote:Originally posted by xyyman: From: Kuwaiti population subgroup of nomadic Bedouin ancestryWhole genome sequence and analysis Sumi Elsa John 1,
quote "273 Kuwaiti natives, we recently demonstrated three distinct genetic subgroups in Kuwaiti population [2]: Kuwait P (KWP) of Persian ancestry; Kuwait S (KWS) of city-dwelling Saudi Arabian tribe ancestry, and Kuwait B (KWB) that includes most of the tent-dwelling Bedouin participants (recruited to provide samples for genotyping). [b]The KWB is distinguished from the other two groups by a characteristic presence of 17% African ancestry (ranging from 11.7% to ***39.4%**)"
---- By African they mean SSA not including North Africans. Include North Africans and ...well...you know the rest
Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
posted
In other words,***ALL*** Bedouins carry SSA(up to 39.4%) and North African ancestry.
Bedouins are Africans just as indigenous North Africans, like Berbers, are Africans. North African extended into Arabia and the Levant. Ottoman Turks infiltrated and displaced the indigenous populations in the Levant.
-------------------- Without data you are just another person with an opinion - Deming Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
Abstract Kuwaiti native population comprises three distinct genetic subgroups of Persian, city-dwelling Saudi Arabian tribe, and nomadic tent-dwelling Bedouin ancestry. Bedouin subgroup is characterized by presence of 17% African ancestry; it owes it origin to nomadic tribes of the deserts of Arabian Peninsula and North Africa.
We sequenced whole genome of a 20 year old male (of Yemeni origin) from the Kuwaiti Bedouin subgroup using Illumina HiSeq 2000. We generated 1273.08 million paired-end reads of length 101 bps that were aligned to the human reference genome hg19. 95.57% of the reads were mapped to the reference genome, resulting in coverage of 41 × (Supplementary Table S1).
Ancestry estimation and haplogroup analysis
Examination of genetic clusters derived using principal component analysis (PCA) for Kuwait population (Fig. 1) reveals that this sample is located deep in the Bedouin cluster (and not at boundaries of the clusters or in regions that overlap among the three clusters). The surname lineage classification identified the participant as a desert-dwelling Bedouin tribe. Further, the ancestry composition of the genetic makeup of the KWB individual is seen as:
European (French_Basque)11%; Arab (Negev Bedouin)44.7%; sub-Saharan African (Biaka_Pygmies)17.3%; and West Asia (Druze, Brahui)24.8%.
This is consistent with the observed compositions in the Bedouin substructure of Kuwaiti population :
European (French_Basque)11%; Arab (Negev Bedouin)45.0%; sub-Saharan African (Biaka_Pygmies)17.0%; and West Asia (Druze, Brahui)25%. As stated in the Introduction,
he nomadic Bedouin subgroup (KWB) is distinguished from the other two groups by a characteristic presence of 17% African ancestry. Arabian ancestry is seen more in the Saudi Arabian tribe ancestry subgroup (KWS) (at 69%) than in the Bedouin group (at 40%); and West Asian ancestry is seen more in the Persian subgroup (KWP) (at 56%) than in any of the other two groups. In order to further illustrate that the ancestry admixture composition of the Bedouin individual sequenced in this study is typical of the KWB group, we present ancestry compositions of 15 samples from the KWS subgroup [10] and one sample from the KWP subgroup [11] (Supplementary Table S2).
While the sequenced Bedouin sample shows presence of 17.3% African ancestry, the 16 samples from the other two subgroups show African ancestry to the extent of only 0.1% to 5.1%; while the sequenced Bedouin sample shows Arabian ancestry at 44.7%; the 15 samples from the KWS subgroup shows Arabian ancestry to a large extent of 66.1% to 86.5%; and while the sequenced Bedouin sample shows only 24.8% West Asian ancestry, the sample from the KWP subgroup shows as high as 64.5%.
The mitochondrial haplogroup (indication of maternal ancestry) of the Bedouin participant is determined as L3d1a1a [L3d], that is predominantly seen in West-Central Africaamong the Fulani [13], Chadians [13], Ethiopians [14], Akan people [15], Mozambique [14], and Yemen [14]. Kivisild et al. [14] analyzed mitochondrial DNA variations in 115 volunteer Yemeni donors in Kuwait (who claimed that their maternal origin was in Yemen) and found that the L macro-haplogroup (the most ancestral mitochondrial lineage) is seen in 47% of the 115 Yemeni individuals; they further found that 20 (17.4%) of the 115 Yemeni participants has the L3 mitochondrial haplogroup (that are most frequently found in sub-Saharan Africa); of these 20 participants, 6 (5.21% of 115 participants) displayed the L3d1 subclade that we observe for the individual sequenced in this study. Thus, the observation of L3d1 haplogroup for the participant in our study is consistent with Yemeni maternal origin. In order to further illustrate that the above observed L3d1 mitochondrial haplogroup is characteristic of the Bedouin sample sequenced in this study, we examined the mitochondrial haplogroups that we identified for a control group of 16 individuals from the other two subgroups of Kuwaiti population (see Supplementary Table S2); none of these 16 samples exhibit the clades of the L macro-haplogroup. Kivisild et al. [14] further compared haplotype diversity seen in Yemeni participants with those reported for Ethiopian population (East Africa); their results highlight the complexity of Ethiopian and Yemeni genetic heritage and are consistent with the introduction of maternal lineages into the South Arabian gene pool from different source populations of East Africa. Horn of Africa (a peninsula in the eastern region of the African sub-continent, enclosing Ethiopia, Somalia, Djibouti and Eritrea) is separated from the south Arabian Peninsula (particularly Yemen) by a short distance of only ~ 10 miles at the strait of Bab-el-Mandeb (the Gate of Tears); the distance across is only ~ 20 miles from Ras Menheli in Yemen to Ras Siyyan in Djibouti. Outside of Africa, L3d is mainly found in African Americans; approximately 6% of all African Americans are descendents of the L3d family line [16].
Posts: 42918 | From: , | Registered: Jan 2010
| IP: Logged |
quote:Originally posted by Brada-Anansi: What about the male line.
The KWB sample is observed to have J1e [J-P58] Y-chromosome haplogroup which is seen in the Arabian Peninsula. The overall estimated time of expansion of J1e haplogroup is around 10,000 years and the ancestors of J1e haplogroups are observed in the Caucasus and eastern Anatolian populations [12]. The frequencies of J1e in populations from the region are [12]: Sudan: (74.2%; n = 35); Yemen (67.7%; n = 62); Negev Bedouin (64.4%; n = 28); Ismaili Damascus (58.8%; n = 51); Qatar (56.9%; n = 72); Jordan (48.7%; n = 76); Sunni Hama (44.4%; n = 36); Oman (37.2%; n = 121); and UAE (34.8%; n = 164). The observed high value of 67.7% for the frequency of J1e in Yemeni population is consistent with the self-reported Yemeni ancestry by the participant.
Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
posted
Eur J Hum Genet. 2010 Mar; 18(3): 348353. Published online 2009 Oct 14. doi: 10.1038/ejhg.2009.166 PMCID: PMC2987219
The emergence of Y-chromosome haplogroup J1e among Arabic-speaking populations
Jacques Chiaroni,1,* Roy J King,2 Natalie M Myres,3 Brenna M Henn,4 Axel Ducourneau,1 Michael J Mitchell,5 Gilles Boetsch,1 Issa Sheikha,6 Alice A Lin,2 Mahnoosh Nik-Ahd,2 Jabeen Ahmad,2 Francesca Lattanzi,7 Rene J Herrera,8 Muntaser E Ibrahim,9 Aaron Brody,10 Ornella Semino,11 Toomas Kivisild,12 and Peter A Underhill2
Abstract
Haplogroup J1 is a prevalent Y-chromosome lineage within the Near East. We report the frequency and YSTR diversity data for its major sub-clade (J1e). The overall expansion time estimated from 453 chromosomes is 10 000 years. Moreover, the previously described J1 (DYS388=13) chromosomes, frequently found in the Caucasus and eastern Anatolian populations, were ancestral to J1e and displayed an expansion time of 9000 years. For J1e, the Zagros/Taurus mountain region displays the highest haplotype diversity, although the J1e frequency increases toward the peripheral Arabian Peninsula. The southerly pattern of decreasing expansion time estimates is consistent with the serial drift and founder effect processes. The first such migration is predicted to have occurred at the onset of the Neolithic, and accordingly J1e parallels the establishment of rain-fed agriculture and semi-nomadic herders throughout the Fertile Crescent. Subsequently, J1e lineages might have been involved in episodes of the expansion of pastoralists into arid habitats coinciding with the spread of Arabic and other Semitic-speaking populations.
Posts: 42918 | From: , | Registered: Jan 2010
| IP: Logged |
posted
J1e males migrating from the Turkey Zagros Mountains and meeting and mating with Sub Saharan African women and they decided to settle in Yemen to have kids and raise a family......
-------------------- Without data you are just another person with an opinion - Deming Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
posted
Keep in mind Africans and Yemen men carry a high frequency of J1e, J1 and J1*. Maybe just maybe the older haplotype of J1e hasn't been found in Africa as yet.......
-------------------- Without data you are just another person with an opinion - Deming Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
quote:Originally posted by Brada-Anansi: What about the male line.
For what it's worth
Genetic structure of the Kuwaiti population revealed by paternal lineages S. Triki-Fendri et al - 2016
quote:...The analyzed markers allowed the discrimination of 19 different haplogroups with a diversity of 0.7713. J-M304 was the most frequent haplogroup in the Kuwaiti population (55.5%) followed by E-M96 (18%). They revealed a genetic homogeneity between the Kuwaiti population and those of the Middle East (FST56.1%, P-value<0.0001), although a significant correlation between genetic and geographic distances was found (r50.41, P-value50.009). Moreover, the nonsignificant pairwise FST genetic distances between the Kuwait population on the one hand and the Arabs of Iran and those of Sudan on the other, corroborate the hypothesis of bidirectional gene flow between Arabia and both Iran and Sudan.
(...) Overall, we have revealed that the Kuwaiti population has experienced significant gene flow from neighboring populations like Saudi Arabia, Iran, and East Africa...
quote:The frequency distribution of haplogroup J exhibits a radial decline from the Middle East toward Central Asia, Caucasus, North Africa, and Europe (Al-Zahery et al., 2011; Cadenas et al., 2008; Quintana- Murci et al., 2001). The Southern Arabian Peninsula has been proposed to be the place of origin of haplogroup J. In fact, the distribution of this haplogroup agrees with the first major expansions proposed fromGMmarker analysis (Chaabani, 2002) and that of mtDNA haplogroup R0a (Cerny et al., 2011) providing evidence for its deep genetic root in Southern Arabia (Chaabani et al., 2000)
Based on the frequencies of both subhaplogroups J1 and J2, the mutation leading to the subclade J1 has, very likely, happened in Yemen (J1 frequency is 72.% vs. 9.6% for J2). By contrast, J2 would have arisen outside of Yemen, particularly in the northern region such as Syria (33.6% for J1 vs. 20% for J2), Turkey (9.1% vs. 24.2%), or Greece (1.9% vs. 18.1%)
-----
On the other hand, haplogroup J2-M172 was observed in 10.2% of the Kuwaiti samples. This haplogroup is highly represented in the Middle East (16% in the population of Saudi Arabia (Abu-Amero et al., 2009), and 23% in the population of Iran (Regueiro et al., 2006), particularly in the Fertile Crescent [25% in the Lebanese population (Zalloua et al., 2008b) and 24% in that of Turkey (Cin- nio? glu et al., 2004)]. In fact, this region has been proposed as the most likely origin of haplogroup J2-M172, which was then spread to Europe during the Neolithic (Cin- nio? glu et al., 2004; Semino et al., 2000, 2004). In
---
In addition, 12% of all of the analyzed Kuwaiti samples carry the haplogroup E-M35 (E1b1a1*). It has been pro- posed that the origin of haplogroup E-M35 could most likely be Eastern Sub-Saharan Africa, where the highest frequencies of this subhaplogroup are observed: 19.2% in the Ethiopian population and 16.7% among the Khoisans of South Africa (Cruciani et al., 2002, 2004; Underhill et al., 2000)....
---
E-M78 carried by 3.4% of the analyzed Kuwaiti individuals, was observed over a wide area, including East Africa (21.5%), North Africa (18.5%), Mid- dle East (5.8%), and Europe (7.2%). The Southern Medi- terranean has been strongly imprinted by this haplogroup (Hammer et al., 1998; Semino et al., 2000, 2004). And the high frequency of E-M78, added to its high microsatellite diversity in Eastern Africa, has led to the hypothesis that it originated from this region, 23.2 ky ago (95% CI 21.1 25.4 ky) (Cruciani et al., 2004, 2007). The cluster Ç, a par- ticular branch of E-M78, defined by the presence of the rare Y-STR allele 11 in DYS19, is observed in Eastern Africa at an average frequency of 17.7% and is almost absent in the populations outside the Horn of Africa (Cru- ciani et al., 2004; Sanchez et al., 2005). In
proposed Phylo tree & a network of weighted Short tandem repeats of the variable Haplogroups found in Kuwait. - from S. Triki-Fendri et al - 2016 (Click to Enlarge) Doi:10.1002/ajhb.22773
In the study above a lil over a hundred unrelated samples were used, however from an unspecified set of locations to my knowledge. nonetheless the results still open up space for postulations or theories such as the following based on dating via BATWING...
quote: Based on time estimation with BATWING, the median TMRCA obtained for haplogroup E-M35 in Ethiopia (12 kya [95% CI 4.550.5]) was almost similar to that of Kuwait (12.8 kya [95% CI 4.151.3]). In fact, it has already been proposed that the likely origin of the subha-plogroup E-M123 could be the Middle East (Cruciani et al., 2004), whereas the second subhaplogroup E-M78 was proposed to be issued from North-East Africa (Cru-ciani et al., 2007). Thus, the similarity between TMRCAs of E-M35 in Kuwait and Ethiopia suggests a bidirectional gene flow between both regions, concordant with historical data on people movements via Bab-el-Mandeb Strait (Chaabani, 2014; Lahr, 1994; Oppenheimer, 2003; Stringer, 2000).
Sited abovePosts: 1781 | From: New York | Registered: Jul 2016
| IP: Logged |
quote: An open question in the history of human migration is the identity of the earliest Eurasian populations that have left contemporary descendants. The Arabian Peninsula was the initial site of the out-of-Africa migrations that occurred between 125,000 and 60,000 yr ago, leading to the hypothesis that the first Eurasian populations were established on the Peninsula and that contemporary indigenous Arabs are direct descendants of these ancient peoples.
This is a trick using wording. The "basal European" populations that appeared on the Arabian Peninsula would have looked no different from the African migrants they separated from. These folks are simply trying to imply that this separation introduced white features into the population which is a characteristic many of these descendant populations carry today as "Eurasians".
Technically these people would still be aptly categorized as African, since there were no "Eurasians" anywhere else yet.
Posts: 8889 | Registered: May 2005
| IP: Logged |
quote: Indigenous Arabs are descendants of the earliest split from ancient Eurasian populations Juan L. Rodriguez-Flores 2016
Odd, so explain what happened?
quote:We use high-resolution genetic data to investigate the genetic and linguistic support for hypotheses concerning the population history in the Chad Basin. The mitochondrial L3f3 haplogroup is found almost exclusively in Chadic speaking populations and its TMRCA corresponds well with archaeological and linguistic dates of the proposed migration of Chadic speaking pastoralists from East or North East Africa to the Chad Basin.
Haplogroup L3f is defined by the coding variants
3396-4218-15514-15944del and the control region motif 1620916519 with a TMRCA of 57,100 ± 9,400 YBP. This haplogroup diversifies into sub-haplogroups L3f1, L3f2 and L3f3. The most geographically widespread sub-haplogroup is L3f1, which is distributed across the African continent [3] and also Arabia [32,33] and has a TMRCA of 48,600 ± 11,500 YBP.
[...]
"The youngest clade, L3f1b2, seems to be more frequent in the Middle East. L3f1a seems to be older (37,700 ± 10,000 YBP) than its sister sub-haplogroup L3f1b and is also less diversified. A few samples from Chad belong to these sub-haplogroups: two to L3f1a and one to L3f1b3."
"We then estimated pairwise FST genetic distances between populations (Additional file 4) and displayed these on a MDS plot (Figure 3). Interesting results are immediately evident while Chadic populations form a relatively homogeneous group, the Cushitic populations split into two completely different clusters. The first group is composed of Horn of African populations, such as Ethiopian and Somali Cushitic populations, which are close to neighbouring Ethiopian Semitic speaking groups and relatively close also to Chadic people from the Chad Basin. The second Cushitic group is composed by more southern groups from Tanzania, i.e. Burunge and Iraqw, who occupy outlier positions even within the Afro-Asiatic MDS plot. In the MDS plot, geography is more strongly associated with genetic distance than is linguistic affiliation.
Overall, we observe that Chadic speaking populations are intermixed with other populations from Chad Basin, including Niger-Congo, Semitic, and Berber speaking people. In this context, it seems that the linguistic categories play a secondary role in structuring the genetic diversity."
--Viktor Černý1 et al.
Migration of Chadic speaking pastoralists within Africa based on population structure of Chad Basin and phylogeography of mitochondrial L3f haplogroup
quote: "Particularly, Yemen has the largest contribution of L lineages (30). So, most probably, this area was the entrance gate of a portion of these lineages in prehistoric times, which participated in the building of the primitive Arabian population."
--Khaled K Abu-Amero et al.
Mitochondrial DNA structure in the Arabian Peninsula
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
Some say that the Mahra people (Arab Salah tribe) (Mehri language speakers) might have a deeper history than others in the region
however the Arabian peninsula has undergone many aridization periods where people may have been migrating in and out
Southern Arabians in general tend to preserve more ancient lineages than northern Arabians but you are correct about populations moving in and out of the area during periods of moisturization and aridization respectively.
Because of such movements, who is to say which Arabs are more indigenous?? What did indigenous Arabs look like?? We all know that modern 'Arab' ethnicity is a branch of Semitic speaking people yet Semitic according ot linguistics was said to have arisen within the last 5,000 years alone.
As Tukuler points out below...
quote:Originally posted by Tukuler: We all know about the Akhdam as their identity and plight was discussed numerous times in this forum. And again, your own "guess" does not matter at all, only the facts do! **Their form of sickle-cell is known as the Arab-Indian HBS and is shared by peoples in both Arabia and India regardless of phenotype** and it is still unknown from which region the allele arose. Also, the Akhdam are not the only 'Negrito' group in Arabia.
The Qarra were also traditionally described as 'Negrito'.
But unlike the Akhdam, the Qarra and Hawt are not marginalized at all but are respected as ancient tribes of the Qahtani or southern division of Arabs. The Akhdam are marginalized primarily because they are considered non-Arab outcasts. The popular theory is that the Akhdam are descended from Ethiopian invaders of the 6th century CE, but the problem with this notion is that there are no archaeological traces tying them to 6th century Ethiopia. Furthermore their physical features are distinct from the Ethiopians/Eritreans across the straits. Even the poster/moderator Yom commented that their beardless faces and very short statures contrast them from the Eritrean peoples. And again, the genetic studies including the Arab-Indian HBS all point to indigenous origins. The likely hood is that the Akhdam represent an indigenous group of people who were not Semiticized/Arabized until relatively recently. They are not the only ones. There are several other groups in Arabia including the Sibyan in the north and certain coastal fishermen who are considered outcasts because their ancestry is considered non-Arab or non-Semitic.
Either way, it becomes evident that these groups didn't exactly look "caucasoid" despite what the Euronuts say.
Posts: 26236 | From: Atlanta, Georgia, USA | Registered: Feb 2005
| IP: Logged |
posted
^ They may not be related to Somalis, but there is genetic evidence that they are related to Habeshas (Abyssinians) of Eritrea and the Ethiopian highlands as shown through Y chromosome hg J1 whose entry in the Horn dates to the Neolithic.
Posts: 26236 | From: Atlanta, Georgia, USA | Registered: Feb 2005
| IP: Logged |
quote:Southeast and south Asian populations are also often thought to be derived from the admixture of various combinations of western Eurasians (Caucasoids), east Asians and Australasians. [ ] These findings, coupled with the recently discovered presence of haplogroup U in Ethiopia [11], support a scenario in which a northeast African population dispersed out of Africa into India, presumably through the Arabian peninsula, before 50,000 years ago (Figure 2). Other migrations into India also occurred, but rarely from western Eurasian populations. [ ] Thus, the caucasoid features of south Asians may best be considered pre-caucasoid that is, part of a diverse north or north-east African gene pool that yielded separate origins for western Eurasian and southern Asian populations over 50,000 years ago.
~Todd R. Disotell. Human evolution: The southern route to Asia Volume 9, Issue 24, 30 December 1999, Pages R925R928
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
posted
^ And you wonder why people still use the term "caucasoid". The word from its inception was already inaccurate and invalid, its been overused and misused for so long it is actually hilarious to see any purported scientific paper use the label.
Posts: 26236 | From: Atlanta, Georgia, USA | Registered: Feb 2005
| IP: Logged |
quote: An open question in the history of human migration is the identity of the earliest Eurasian populations that have left contemporary descendants. The Arabian Peninsula was the initial site of the out-of-Africa migrations that occurred between 125,000 and 60,000 yr ago, leading to the hypothesis that the first Eurasian populations were established on the Peninsula and that contemporary indigenous Arabs are direct descendants of these ancient peoples.
This is a trick using wording. The "basal European" populations that appeared on the Arabian Peninsula would have looked no different from the African migrants they separated from. These folks are simply trying to imply that this separation introduced white features into the population which is a characteristic many of these descendant populations carry today as "Eurasians".
Technically these people would still be aptly categorized as African, since there were no "Eurasians" anywhere else yet.
Most likely the "basal Eurasians" or would be "backflowees" woud have looked like Africans..
XYZMAN says: They are after basal Eurasian and the Neolithics. I told you this is where the new war is at. They have ceded AEians to Africans now they bendng over backwards to wrestle Neolithics from Africa. They dont want to be classified as subset of Africans. Ooops! Too late. Lol!
lol.. even with the "basal" thing they are still a subset...
Posts: 5905 | From: The Hammer | Registered: Aug 2008
| IP: Logged |
posted
^ I disagree with xyzman-- mainstream scholarship has not yet "ceded" ancient Egypt to Africans at least not yet. From what I've been reading in many science and history websites they have only just begun the process of accepting this undeniable reality. However, I do agree that the big battle now centers around the forebears of Neolithic culture in the Middle East, and the Lazaridis et al. findings of "basal Eurasian" have only instensified the battle.
According to the simplified graph above, 'Non-Africans' that is OOA ancestors share a common ancestry with Mota boy of Ethiopia. It is then that OOA splits into 'basal Eurasians' and 'main Eurasians'. This brings us back to the question of where exactly the 'Non-African' or OOA ancestors diverged. Did they diverge in Southwest Asia or still in the African continent? According to Swenet, he thinks OOA diverged in Africa and even after the first major OOA expansion after the Toba event, there were still OOA populations left in Africa. If this is the case then 'Non-African' or OOA should be labeled as something else. Even the Y-chromosomal label of "Eurasian Adam" for example if misleading because many geneticsts agree that this ancestor arose in Africa first before leaving. But definitely the 'basal Eurasians' must have been of recent African extraction due to the peculiarly African even "negroid" traits found associated with their fossils as seen in not only some Natufian remains but the remains of their alleged ancestors-- the Kebarans. This only proves that Southwest Asia was indeed an extension of Africa more than most people realize.
Posts: 26236 | From: Atlanta, Georgia, USA | Registered: Feb 2005
| IP: Logged |






 UBBFriend: Email this page to someone!
UBBFriend: Email this page to someone!

























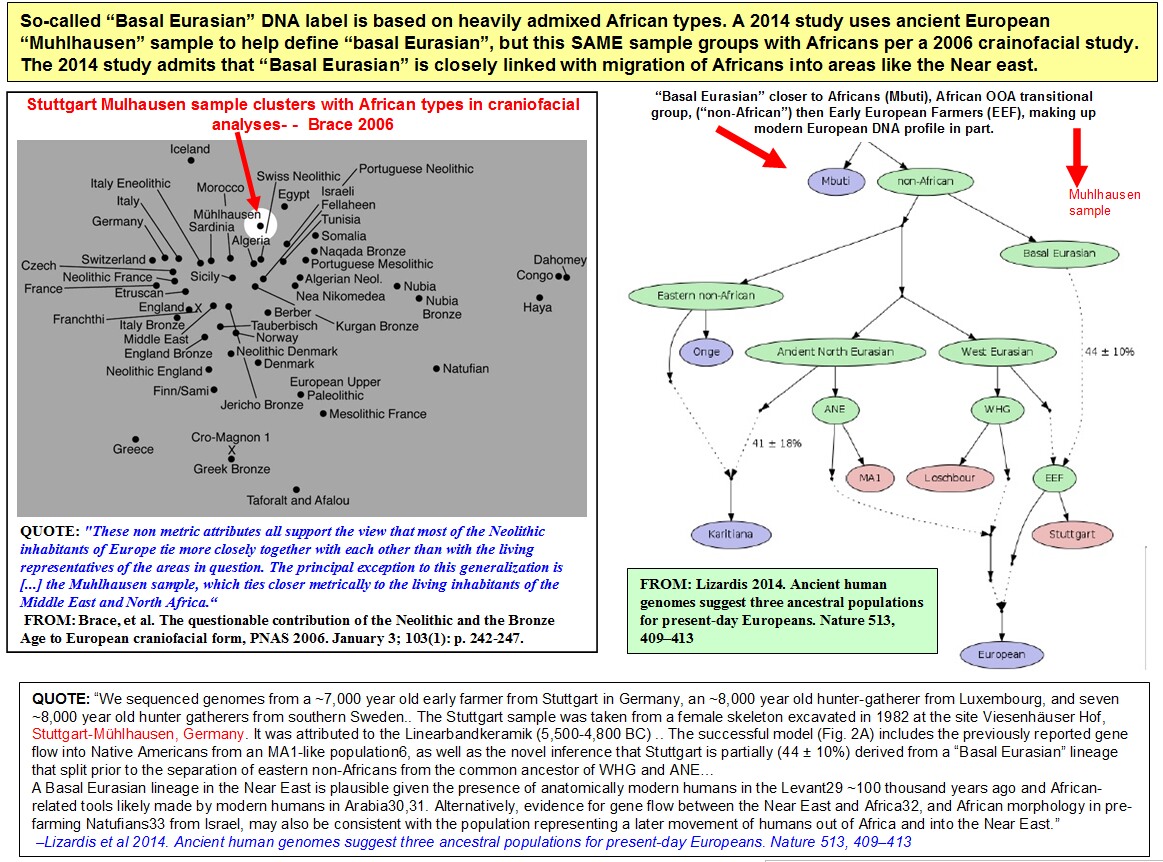





 Printer-friendly view of this topic
Printer-friendly view of this topic