Were Rivers Flowing across the Sahara During the Last Interglacial? Implications for Human Migration through Africa2013 Tom J. Coulthard mail,
Jorge A. Ramirez, Nick Barton, Mike Rogerson, Tim Brücher
excerpts
Abstract
Human migration north through Africa is contentious. This paper uses a novel palaeohydrological and hydraulic modelling approach to test the hypothesis that under wetter climates c.100,000 years ago major river systems ran north across the Sahara to the Mediterranean, creating viable migration routes. We confirm that three of these now buried palaeo river systems could have been active at the key time of human migration across the Sahara. Unexpectedly, it is the most western of these three rivers, the Irharhar river, that represents the most likely route for human migration. The Irharhar river flows directly south to north, uniquely linking the mountain areas experiencing monsoon climates at these times to temperate Mediterranean environments where food and resources would have been abundant. The findings have major implications for our understanding of how humans migrated north through Africa, for the first time providing a quantitative perspective on the probabilities that these routes were viable for human habitation at these times.
Introduction
The role of the Sahara as a geographical filter and launch zone for dispersals of Homo sapiens out of Africa is a controversial topic [1], [2], [3], [4]. At issue is the observation that 130-100,000 years ago there was a marked increase in humidity in the present desert and adjacent regions [5], [6], [7], [8], which coincided with some of the earliest appearances of H. sapiens in both the Sahara and the Levant [9], [10], [11], [12], [13], [14], [15], [16]. During MIS 5 (Marine Isotope Stage 5) [17], high insolation in the northern hemisphere caused the African monsoon to assume a position up to 1,000 km north of its location today [18], [19], [20], [21]. Isotopic and geomorphic evidence suggests that rain falling on the north of the Trans-Saharan mountains then flowed towards the Mediterranean [2], [22], [23], potentially creating migration pathways via a series of green corridors [4] and mega lake corridors [24] across the Sahara. Dating of human fossils from the Levantine sites of Skhul and Qafzeh imply that early dispersals occurred along the eastern margins of the Sahara prior to ~100 ka [13], [14], [15], while other craniodental remains show that populations closely resembling those of the Near East were simultaneously present in north western Africa [9], [16], [18]. Given the combined dating uncertainties, many of these fossils and associated archaeological Middle Palaeolithic/Middle Stone Age finds are likely contemporary with the last interglacial period of peak humidity attested in marine cores [20], [23] and stalagmites [6], [25].
However, supporting evidence for the green corridor hypothesis remains subjective. Aside from topographic analysis, there has been no quantitative hydrological assessment (i.e. calculation of fluxes and balances of water) that could test whether these freshwater pathways across the Sahara were physically possible during the Eemian (MIS 5e). Surface evidence of fossil river systems and dated lacustrine records show there has been water in the region, but this does not provide an effective quantitative view of when, where, or how much water was present in the wider landscape. Interpretation of the archaeological record is equally subjective, as although the general direction of dispersal (northward from sub-Saharan Africa) and its general timing (last interglacial) are clear, these data remain ambiguous between a single trans-Saharan migration with delayed subsequent expansion, multiple migrations via a single route or multiple migrations via multiple routes. The existing evidence is not sufficient to conclude whether contiguous Green Corridors existed at the right time for migration.
In this paper, for the first time, we simulate the balance and fluxes of water across this region. We use simulated precipitation from a state of the art Earth System Model (ESM) simulation of the Eemian (MIS 5e) climate to drive a combined hydrological and hydraulic model to reconstruct past rivers and flood events across 12,000,000 km2 of North Africa. For the first time, this reveals the seasonal and spatial patterns of Saharan surface palaeohydrology, predicting the presence of distinct river corridors and wetlands [24], [26]. Our simulations were carried out with the sole aim of testing the Green Corridor hypothesis; were contiguous corridors following surface water (i.e. rivers) really feasible during MIS 5e? Do all the buried rivers show the same history, or are there spatiotemporal differences? We achieve this by calculating a probability of surface water routes across the Sahara as a basis for further investigation.
Results
Figure 2 demonstrates the existence of a series of extensive ephemeral and perennial river systems draining North from the Ahaggar and Tibesti mountains across the Sahara to the Mediterranean during the period 125124 ka BP. Some channels dissipate in the desert, but some converge forming three main systems; in the West the Irharhar river draining into the Chott Melrir basin, and to the East two larger systems named the Sahabi and the Kufrah (Fig. 2). As the rainfall is associated with the monsoon, flow is highly seasonal and the Irharhar river is ephemeral, flowing for ~3 months. The Sahabi and Kufrah systems are close to perennial, due to larger contributing areas in the catchment headwaters located in the monsoon belt (Fig. 1). Figure 3 demonstrates the seasonality of flow in all three systems with precipitation in August taking over two months to reach the coast or near-coastal lake systems. The distinct climatic zoning (Fig. 1) means all three rivers are allogenic, losing water along their length with little or no hydrological contribution once they leave the mountains/uplands. In addition to rivers, the simulations predict massive lagoons and wetlands in NE Libya some of which are extensive (>70,000 km2). These are also fed from the Jebel Akhdar in Cyrenaica, which also received higher rainfall during this period. There are smaller lakes forming in Tunisia and Algeria due to water supplied via the Irharhar river system.
Discussion
This study provides the first strong quantitative evidence for the presence of three major river systems flowing across the Sahara during MIS 5e. We simulated three river systems that are now largely buried by dune systems, but when flowing would have provided fertile habitats for flora and fauna in proximity to the channels [22]. Notably, the Sahabi and Kufrah would be major river systems with monsoon discharges significantly in excess of 2500 m3 s−1 and an extensive system of anabranches and wetlands. In the Libyan Kalanschiou region, the green corridor would have been 100 km wide, substantial and largely perennial. This reconstruction is highly compatible with evidence of widespread palaeosols deposited on the margins of this system during the less pronounced Holocene humid period [22]. Here we have simulated one wet phase, but this research strongly supports the occurrence of similar Green Saharas recorded in the marine [37] and terrestrial [26] archive.
Our simulation results quantitatively confirm previous hypotheses of these rivers shown in geomorphic and geochemical data [2], [4], [22], [23], [24]. For example, the radio-isotopic composition of the water identified in the Ionian Sea [4], [38] indicates that it was precipitated in the basaltic trans-Saharan range in Southern Libya as shown by our results. Runoff flowing rapidly to the coast, in a manner highly comparable to our simulated runoff waves, is shown by the light oxygen isotopic composition of the water flowing into the Mediterranean at this time [20]. Furthermore, the river systems that our research simulates are consistent with the well-preserved drainage network that has been identified in these regions by fieldwork and from satellite imagery [22], [24]. Overall, our confirmation that these hypotheses are physically realistic allows us to move on to questions of how and when the rivers operated, rather than their existence.
Whilst we cannot state for certain that humans migrated alongside these rivers, the shape of the drainage systems indicate that anyone moving from south to north from a 2000 km wide region in the mountains would be funnelled into three clear routes. There is also a clear geographical split, with a 2000 km gap between the destinations of Irharhar and the combined Sahabi and Kufrah systems.
Despite being ephemeral, the Irharhar river corridor could be the most suitable for dispersal of hominids beyond the Sahara. Uniquely, the Irharhar extends from humid to humid climes - ranging from the monsoonal Ahaggar and Tibesti region to the North Western Mediterranean climate zone that also received substantial winter rainfall (Fig. 1). High humidity in the destination region during the last interglacial is confirmed by the presence of significant water near the Chott Melrir basin [39]. Whilst the more extensive Sahabi and Kufrah also traverse the Sahara, their downstream limits remain within the arid/semi-arid regions [40].
Support for the significance of the Irharhar river corridor is provided by the high number of Middle Stone Age archaeological sites concentrated in the western region (Fig. 2). Many of these locations contain Levallois lithic artefacts with Aterian affinities that on comparative grounds can be plausibly dated to the last Interglacial [41], [42]. It is highly likely given the existing artefact distributions that humans migrated northwards from the relatively humid Trans-Saharan mountainous zones to the Maghrebian Mediterranean biome (Fig. 2). The loose clustering of sites along our simulated Irharhar river and associated channels implies this as a preferred route of dispersal. Furthermore, as the simulations are driven by present day topography, if the dune systems in this region were removed or reconfigured the Irharhar could flow further to the West. In contrast, the eastern region has a surprising lack of archaeological evidence despite the extensive simulated palaeo-river courses. It is likely that further surveys in this area will provide substantial evidence of Middle Stone Age activity, especially in the areas of buried palaeochannels. However, continued absence of this critical evidence of human migration would confirm our suggestion that a key factor in the western distribution of sites was the attractiveness of the richer Mediterranean-type environments of the Maghreb, which would have promoted permanent settlement in the region and further transit in both directions along the Irharhar river corridor.
Conclusions
For the first time, our simulations demonstrate that Saharan Humid Corridors were highly likely during the last interglacial strongly re-affirming the viability of these routes as migratory corridors for early hominids. This research provides an unprecedented means of developing new hypotheses for past human, faunal and floral activity in this region and for validating the performance of palaeo climate simulations
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
Nick A. Drakea,1, Roger M. Blenchb, Simon J. Armitagec, Charlie S. Bristowd, and Kevin H. Whitee
a Department of Geography, Kings College London, Strand, London WC2R 2LS, United Kingdom; b Kay Williamson Educational Foundation, 8 Guest Road, Cambridge CB1 2AL, United Kingdom; c Department of Geography, Royal Holloway, University of London, Egham, Surrey TW20 0EX, United Kingdom; dSchool of Earth Sciences, Birkbeck College, University of London, Malet Street, London WC1E 7HX, United Kingdom; and eDepartment of Geography, University of Reading, Whiteknights, Reading RG6 6AB, United Kingdom
Edited by Ofer Bar-Yosef, Harvard University, Cambridge, MA, and approved November 22, 2010 (received for review August 23, 2010)
Ancient watercourses and biogeography of the Sahara explain the peopling of the desert
Evidence increasingly suggests that sub-Saharan Africa is at the center of human evolution and understanding routes of dispersal out of Africa is thus becoming increasingly important. The Sahara Desert is considered by many to be an obstacle to these dispersals and a Nile corridor route has been proposed to cross it. Here we provide evidence that the Sahara was not an effective barrier and indicate how both animals and humans populated it during past humid phases. Analysis of the zoogeography of the Sahara shows that more animals crossed via this route than used the Nile corridor. Furthermore, many of these species are aquatic. This dis- persal was possible because during the Holocene humid period the region contained a series of linked lakes, rivers, and inland deltas comprising a large interlinked waterway, channeling water and an- imals into and across the Sahara, thus facilitating these dispersals. This system was last active in the early Holocene when many spe- cies appear to have occupied the entire Sahara. However, species that require deep water did not reach northern regions because of weak hydrological connections. Human dispersals were influenced by this distribution; Nilo-Saharan speakers hunting aquatic fauna with barbed bone points occupied the southern Sahara, while peo- ple hunting Savannah fauna with the bow and arrow spread south- ward. The dating of lacustrine sediments show that the green Sahara also existed during the last interglacial (∼125 ka) and pro- vided green corridors that could have formed dispersal routes at a likely time for the migration of modern humans out of Africa.
posted
Domestication Processes and Morphological Change Through the Lens of the Donkey and African Pastoralism Fiona Marshall and Lior Weissbrod
Fiona Marshall is Professor in the Department of Anthropology, Washington University, Saint Louis (1 Brooking Drive, Saint Louis, Missouri 63130, U.S.A. [fmarshal@artsci.wustl.edu]). Lior Weissbrod is a postdoctoral researcher at the Zinman Institute of Archaeology, University of Haifa (Mount Carmel, Haifa 31905, Israel [lweissbr@research.haifa.ac.il]).
Little is known about the beginnings and spread of food production in the tropics, but recent research suggests that definitions that depend on morphological change may hamper recognition of early farming in these regions. The earliest form of food production in Africa developed in arid tropical grasslands. Animals were the earliest domesticates, and the mobility of early herders shaped the development of social and economic systems. Genetic data indicate that cattle were domesticated in North Africa and suggest domestication of two different African wild asses, in the Sahara and in the Horn. Cowpeas and pearl millet were domesticated several thousand years later, but some intensively used African plants have never undergone morphological change. Morphological, genetic, ethnoarchaeological, and behavioral research reveals relationships between management, animal behavior, selection, and domestication of the donkey. Donkeys eventually showed phenotypic and morphological changes distinctive of domestication, but the process was slow. This African research on domestication of the donkey and the development of pastoralism raises questions regarding how we conceptualize hunter-gatherer versus food-producer land use. It also suggests that we should focus more intently on the methods used to recognize management, agropastoral systems, and domestication events.
This paper was submitted 13 XI 09, accepted 02 XII 10, and electronically published 08 VI 11.
The question of whether understanding of the beginnings of food production is being constrained by definitions and methods of detection that focus on morphological change rather than management is becoming a major theme in studies of the origins of agriculture. Recent research in the humid tropics of southeastern Asia and the Pacific suggests that definitions that depend on morphological change hamper recognition of early farming in these areas (Bayliss-Smith 2007; Denham 2007, 2011). This perspective has so far centered on plants of the humid tropics that have a history of long-term cultivation in agricultural systems but lack morphological change (Denham 2007; Kahlheber and Neumann 2007; Yen 1989). Another feature of both humid and arid tropical agricultural practices that has strained conceptions of early agricultural systems is the variety of economic activitiesincluding fishing, gathering, hunting, cultivation, and herdingthat may be combined in complex and diverse subsistence systems (Kahlheber and Neumann 2007; Marshall and Hildebrand 2002; for North America, Smith 2001, 2011).
In their approach to definitions and the question of whether morphological change is an effective marker of domestication, Jones and Brown (2007) focus on selection processes and timing rather than on region. They contend that under certain circumstances, practices of cultivation and protective tending could have resulted in stable long-term systems of food production that depended on plants and animals lacking distinctively domestic morphological and genetic characteristics. Reproductive isolation and morphological change, Jones and Brown (2007) go on to suggest, are linked with later stages of agricultural development, when human populations expanded and people removed plants and animals from their wild ranges.
There is a growing appreciation, however, of differences among species in time elapsed before domestication processes are readily detectable and of variability in the sensitivity of methods that can be brought to bear on any given taxon. In a detailed study of the domestication of goats in western Asia, Zeder (Zeder 2008; Zeder and Hesse 2000) used regional and age- and sex-based variability in animal size to document early herd management, which was followed by diminution in size. In the absence of clear morphological indicators, evidence for managementculling, corralling, and milkinghas also been key to a better understanding of early phases of domestication of the horse (Outram et al. 2009). The discovery by Rossel et al. (2008) that donkeys used by Egyptian pharaohs for transport at approximately 5000 cal BP (historic date 3000 BC; table 1) remained morphologically wild 1,000 years after they were thought to have been first domesticated further emphasizes possibilities for underestimating the timing of domestication of large mammals and draws attention to species-specific pathways to domestication (see also Zeder 2011).
Table 1. Key African animal and plant domesticates, with summaries of sites, date ranges, and arguments for management or domestication processes
In the light of these different emphases on global, regional, and taxon-specific impacts of late morphological change on general understanding of early food production, we evaluate current perspectives on the beginnings of food production in Africa, a continent that represents the worlds largest tropical landmass. We reexamine evidence of early animal and plant domesticates and employ ethnoarchaeological data on donkey management and breeding behavior to examine species-specific domesticatory practices that influenced selection and the likelihood of morphological change. These analyses allow us to return to the larger question of Africas contribution to understanding variability in early agricultural systems worldwide. In most of Africa, pastoralism is considered the earliest form of agriculture, followed by plant cultivation and adoption of mixed herding-cultivation systems.
Early Food Production in AfricaJump To Section...
Africanists have built up a picture of the beginnings of food production in which early dependence on domestic animals and increasing reliance on mobility guided the development of social and economic systems of the Early Holocene and resulted in late domestication of African plants. Specific themes that have emerged include locally and socially contingent responses to large-scale climatic change, domestication of cattle for food and donkeys for transport, intensive hunting and possible management of Barbary sheep, long-term reliance on a broad range of wild plants and animals, and late domestication of African plants.
In this review of the African evidence, we see domestication as a microevolutionary process that transformed animal and plant communities and human societies (see Clutton-Brock 1992), but we examine rather than assume relationships between domestication and long-term genetic and morphological change (see also Vigne et al. 2011). We follow Zeder (2009, 2011; Rindos 1984) in emphasizing long-term coevolutionary relations between people, animals, and plants, but unlike Rindos (1984), we also highlight the intentional role that individuals played in selection (Hildebrand 2003b; Marshall and Hildebrand 2002). Pastoralism is also an important concept for discussions of the beginnings of food production in Africa, and this, we argue, differs from herding or simple keeping of animals because pastoralists rely on moving livestock to pasture and emphasize the social and symbolic role of domestic animals (Dyson-Hudson and Dyson-Hudson 1980; Smith 2005; Spear and Waller 1993). This does not necessarily imply, however, a diet heavily based on domestic animals. Historically, African pastoralists prioritized the needs of their herds in scheduling activities and locating settlements (McCabe 2004; Western and Dunne 1979), but they usually relied on a broad range of complementary subsistence strategies ranging from seasonal cultivation, fishing, hunting, and gathering to food exchange or trade (Dyson-Hudson and Dyson-Hudson 1980; Evans-Pritchard 1940; Schneider 1979). As a result, it is overly simplistic to rely on high proportions of domestic animal bones to differentiate pastoral from hunter-gatherer or farming sites. Multiple lines of evidence are necessary, including households oriented to mobilitywith slope, soil, and vegetation characteristics organized around the needs of domestic herds (Western and Dunne 1979)animal pens, dung deposits (Shahack-Gross, Marshall, and Weiner 2003; Shahack-Gross, Simons, and Ambrose 2008), milk residues (see Evershed et al. 2008), livestock-focused rock art, and ritual livestock burials (di Lernia 2006).
Domesticatory Settings: Climatic and Social Variability and Subsistence Intensification
Large-scale climate change forms the backdrop to the beginnings of food production in northeastern Africa (Kröpelin et al. 2008). Hunter-gatherer communities deserted most of the northern interior of the continent during the arid glacial maximum and took refuge along the North African coast, the Nile Valley, and the southern fringes of the Sahara (Barich and Garcea 2008; Garcea 2006; Kuper and Kröpelin 2006). During the subsequent Early Holocene African humid phase, from the mid-eleventh to the early ninth millennium cal BP, ceramic-using hunter-gatherers took advantage of more favorable savanna conditions to resettle much of northeastern Africa (Holl 2005; Kuper and Kröpelin 2006). Evidence of domestic animals first appeared in sites in the Western Desert of Egypt, the Khartoum region of the Nile, northern Niger, the Acacus Mountains of Libya, and Wadi Howar (Garcea 2004, 2006; Pöllath and Peters 2007; fig. 1).
During the Early and mid-Holocene, diverse hunter-gatherer groups lived close to permanent water in widely separated regions of northeastern Africa, from the Acacus to Lake Victoria (Caneva 1988; Garcea 2006; Holl 2005; Prendergast and Lane 2010). Ethnoarchaeological research suggests that this social and economic variability played a significant role in pathways to food production in Africa. Recent hunter-gatherers with long-term investment in hive and trap construction and delayed-return social systems and limited sharing have historically been able to accommodate more easily property-rights issues arising out of time investment in agriculture than have those with highly egalitarian norms (Brooks, Gelburd, and Yellen 1984; Dale, Marshall, and Pilgram 2004; Marshall 2000; Smith 1998; Woodburn 1982). Moreover, cattle herding requires significantly greater commitment than cultivation because foragers can tend crops intermittently and accommodate them into flexible hunter-gatherer schedules, whereas animal herds require protection against predators and constant attention (Dale, Marshall, and Pilgram 2004; Marshall 2000). As a result, Africanists have hypothesized that domestication of cattle is more likely to have been undertaken and pastoralism adopted in regions of northeastern Africa that were occupied by complex rather than highly mobile egalitarian hunter-gatherers (Marshall and Hildebrand 2002).
Arguments that complex or delayed-return systems of social organization existed in the Acacus, the Sudanese Nile Valley, and some other regions of the African Early to mid-Holocene are based on elaboration of material culture, including manufacture of ceramics and storage facilities in these areas and highly patterned use of rock-shelter sites and local landscapes (Barich 1987; di Lernia 1999, 2001; Garcea 2004; McDonald 2008). Significant investment in living spaces and limited movement are indicated by hut construction at Nabta Playa in the Acacus Mountains and the northern Sudanese Nile Valley and by isotopic analyses at Gobero in Niger and Acacus sites (Barich 1987; Garcea 2006; Sereno et al. 2008; Tafuri et al. 2006). In the central Sahara, the Sudanese Nile Valley, and the Acacus, human burials are common (Caneva 1988; Honegger 2004; Sereno et al. 2008). Garcea (2004) and di Lernia (1999, 2001) argue that their presence in the Late Acacus phase (ca. 10,2509600 to 98909440 cal BP) may relate to group identities and rights to land.
North African hunter-gatherers of the Early and mid-Holocene employed highly diverse subsistence as well as social systems. Wild cattle (Bos primigenius) were hunted along the Mediterranean coast and the Nile Valley, and small numbers of wild ass (Equus africanus) were also present in many sites (Alhaique and Marshall 2009; Gautier 1987a; Marshall 2007). Barbary sheep (Ammotragus lervia) were the most common animal hunted across North Africa at this time (di Lernia 2001; Gautier 1987a; Saxon et al. 1974). In the Late Acacus sites of Ti-n-Torah, Uan Tabu, and Uan Afuda, intensive exploitation of wild cereals (e.g., Echinochloa, Panicum, Setaria, Digitaria, and Pennisetum) is associated with heavy grindstone use (di Lernia 1999; Garcea 2001; Mercuri 2001; fig. 1). A similar set of wild grass seeds were harvested, processed, and stored in the eastern Sahara during the late tenth and early ninth millennia at Nabta Playa, site E-75-6 (Wasylikowa et al. 1993; Wendorf and Schild 1998; for radiocarbon dates, see table 2). Along the Sudanese Nile, a variety of wild mammals were hunted in conjunction with fishing for large deepwater fish and intensive grindstone use (Caneva 1988; Haaland 1987).
There has been a recurrent suggestion that some North Africans penned and culled Barbary sheep herds during early phases of the Holocene (di Lernia 1998, 2001; Garcea 2006; Saxon et al. 1974; table 1). Earlier arguments for management without morphological change were based on young maledominated culling profiles from the sites of Tamar Hat and Haua Fteah on the Mediterranean coast (Saxon et al. 1974; Smith 2008; fig. 1). More recent evidence is based on the presence of dung accumulations in the rear of rock-shelter sites occupied by complex hunter-gatherers during the tenth and early ninth millennia cal BP in the Libyan Acacus at Uan Afuda, Uan Tabu, and Fozzigiaren (Cremaschi and Trombino 2001; di Lernia 2001; Garcea 2006). Di Lernia (2001) argues that dense dung deposits in these rock shelters differ from natural dung accumulations characterized by loose and scattered pellet matrices and result instead from use of shelters for corralling animals. Micromorphological analyses of the dung layer sediments suggest trampling and indicate the presence of spherulites common in caprine dung, and studies of the plant remains indicate a selected range of plant species suggestive of foddering (Castelletti et al. 1999; di Lernia 2001; Mercuri 1999). Interestingly, Livingstone Smith (2001) notes that hunter-gatherer pottery of Late Acacus levels at Uan Afuda is dung tempered, a characteristic of later pastoral ceramics. The number of Barbary sheep remains declines in later sites, however, and there are no dung deposits that suggest subsequent emphasis on Barbary sheep (di Lernia 1999; Garcea 2001, 2004). Taken together, the micromorphological and archaeological evidence for dung accumulation resulting from penning of Barbary sheep in the Late Acacus rock shelters is suggestive, but additional faunal data and dung deposits are needed from open-air sites.
Domestication of African cattle?
The evidence for taming of wild cattle during the Early Holocene provides an interesting parallel to that for management of Barbary sheep. Wendorf and colleagues (Gautier 1987b; Wendorf and Królik 2001; Wendorf and Schild 1998; Wendorf, Schild, and Close 1984) have argued that seasonally settled hunter-gatherers of the Nabta Playa region (fig. 1) domesticated African cattle in the Western Desert of Egypt during the eleventh to tenth millennium cal BP (reviews of arguments in Gifford-Gonzalez 2005; table 2). Domestic sheep and goats, on the other hand, were introduced to Africa from southwestern Asia during the early eighth millennium cal BP and postdate the appearance of cattle at all sites except Uan Muhaggiag (Gautier 2001; Linseele 2010; Linseele et al. 2010). The independent domestication of African cattle has been tied to arid episodes, the desire of hunter-gatherers for increased short-term predictability in food resources, and the difficulty of intensifying plant foods under these conditions (Marshall and Hildebrand 2002). Bos remains are ubiquitous in sites of the Nabta and Bir Kiseiba regions (fig. 1) from the eleventh to the tenth millennium cal BP (table 2) but in very small numbers, precluding detailed analyses of morphometric change or reconstruction of culling profiles (Gautier 2001). Linseele (2004) has demonstrated, however, that size decrease is not a useful indicator of domestication in northeastern Africa because the size of African Bos primigenius varied regionally and temporally and because ancient Egyptian longhorn cattle overlapped in size with some wild cattle populations.
Close and Wendorf (1992) and Gautier (1984b, 1987b) also argued, largely on the basis of a well and a watering basin at site E-75-6, that the repeated presence of water-dependent North African B. primigenius in Western Desert sites during the tenth and ninth millennia cal BP (table 2) reflected range extension facilitated by management and watering of cattle (table 1). Bos cranial remains in a human grave at El Barga in northern Sudan further support the presence of cattle in the region during the early ninth millennium cal BP (Honegger 2005:247248). The earliest evidence of small domestic cattle from the central Sahara dates, however, to the eighth millennium BP (at Ti-n-Torha and Uan Muhaggiag; Gautier 1987b; fig. 1).
To date, the strongest evidence for domestication of cattle in Africa comes from a series of major studies of the genetic characteristics and biodiversity of contemporary cattle breeds. Changing genetic approaches are reviewed by Larson (2011). Initial analyses of maternal mitochondrial DNA (mtDNA) showed that African cattle shared a distinctively higher frequency of the T1 mitochondrial haplogroup than is common in other regions and a large proportion of unique haplotypes (Bradley et al. 1996). These findings are consistent with an independent African domestication, although the possibility of a demographic expansion of Near Eastern cattle in Africa could not be ruled out (Bradley and Magee 2006; but see Achilli et al. 2008). Recent analysis of single-nucleotide polymorphisms from whole-genome sequences derived from small numbers of cattle demonstrate that African breeds diverged early from the European taurine cattle (Decker et al. 2009). New analyses of high-resolution interspersed multilocus microsatellites on the male-specific region of the Y chromosome demonstrate the existence of an African subfamily in taurine cattle of the Y2 haplogroup (Pérez-Pardal et al. 2009). Associated analyses also indicate that neither the genetic diversity in the African mtDNA T1 haplogroup nor the diversity in the Y2 haplogroup is consistent with the bottleneck that would have been required to fix these haplotypes from Near Eastern taurine cattle (Pérez-Pardal et al. 2010; see also Bovine HapMap Consortium 2009). Taken together with data on variation in autosomal microsatellites (rapidly evolving regions of the nuclear genome) and other data on Y-chromosome variability in African cattle breeds (Bradley and Magee 2006; Hanotte et al. 2002), the genetic data as a whole point strongly to an independent African domestication of cattle (Pérez-Pardal et al. 2009, 2010).
Ethnographic studies suggest, however, that genetic and phenotypic change may have been slow in early northeastern-African cattle and that neither morphological nor genetic studies are likely to detect the early phases of this process. Given recurrent cycles of drought and disease, contemporary African pastoralists manage their herds for maximum growth by keeping a high proportion of females in herds (Dahl and Hjort 1976). However, the main intentional selective processes acting on African cattle are culling and castration, which affect males rather than females (Dahl and Hjort 1976; Ryan et al. 2000). Natural selection in the form of drought and disease often play a larger role in mortality than culling (Mutundu 2005), multiple bulls are common in herds, offtake is low (4%8%), and culling often takes place after sexual maturity (Ryan et al. 2000). Such processes, together with some introgression with wild bulls, are likely to have worked against rapid morphological change in early pastoral herds and to have resulted in a postmanagement lag in morphological change.
Domestication of the donkey.
It has long been suggested that ancient Egyptians domesticated the donkey (Equus asinus), although the Near East has also been considered a possible area of origin. Egyptian Predynastic sites have yielded the earliest potential domestic donkeys, which date to the mid-seventh millennium cal BP (historic date 46004400 BC; Boessneck and von den Driesch 1990; table 1). Some faunal elements from these sites, zooarchaeologists argue, exhibit size decrease relative to the wild ass (Boessneck and von den Driesch 1990), but widespread morphological change was slow to develop in ancient Egypt. Evidence of bone pathologies from early dynastic donkey burials at Abydos (fig. 1) demonstrates that by approximately 5000 cal BP (historic date 3000 BC), First Dynasty Egyptian kings were using donkeys to carry heavy loads (Rossel et al. 2008). Rossel et al. (2008) show, however, that these animals were not yet morphologically distinguishable from the African wild ass.
Recent studies of genetic variability in modern donkeys suggest that prehistoric pastoralists may have domesticated donkeys on the fringes of the Sahara. Beja-Pereira and colleagues (2004; also Vilá, Leonard, and Beja-Pereira 2006) document the existence of two different haplogroups or clades of domestic donkeys. Their genetic-diversity data suggest two domestication events, both in northeastern Africa. Kimura et al.s (2010) recent analysis of ancient DNA from the Nubian donkey (Equus africanus africanus) and the Somali wild ass (Equus africanus somaliensis) demonstrates that the Nubian wild ass was the ancestor of modern donkey Clade I but that the ancestor of donkeys of Clade II is currently unknown. This research also documents the ancient distribution of the Nubian wild ass and Clade I donkeys from the Atbara River and Red Sea Hills in Sudan and northern Eritrea across the Sahara to Libya, a geographic distribution that suggests that prehistoric pastoralists domesticated Clade I donkeys (Kimura et al. 2010). However, domestication by pastoralists or farmers of the northern Nile Valley during late prehistoric/early Predynastic times is also a possibility.
The Herding-Hunting Mosaic and the Spread of Pastoralism
In the central Sahara, cattle became common in the eighth to sixth millennium cal BP at sites such as Ti-n-Torha, Uan Muhaggiag, Uan Telocat, Adrar Bous, Gobero, Enneri Bardagué, and Wadi Howar (Clark et al. 2008; di Lernia 2006; Garcea 2004; Gautier 1987b; Jesse et al. 2007; Roset 1987; Sereno et al. 2008; fig. 1). The main advantages for hunter-gatherers of herding cattle over intensification of plant resources or reliance on hunting and gathering are thought to have been decreased reliance on local rainfall and increased predictability in daily access to cattle herds for blood, meat, and ceremonial purposes (Jesse et al. 2007; Marshall and Hildebrand 2002). Foraging continued, but the intensity of the new human-animal relationship would have required ownership patterns and schedules oriented to animal care and transformation of hunter-gatherer societies. Dependence on wild calories could have been somewhat reduced, however, by milking, a practice that archaeologists have tended to assume was adopted after herding for blood and meat and with some difficulty (but see Linseele 2010).
Different genetic bases for lactase persistence in Europe and Africa show coevolution between people and cattle and the strong selective advantage conferred by drinking milk (Tishkoff et al. 2006). Interestingly, recent research has documented lactase persistence among some contemporary African hunter-gatherers. Tishkoff et al. (2006, supplementary information) note that lactase persistence could be selected for by delaying weaning of infants and, moreover, that the trait is also adaptive for digestion of certain roots and barks. This suggests several pathways to lactase persistence among hunter-gatherers and raises the question of whether African herders milked their cattle earlier and incorporated dairy products into their diets with fewer digestive difficulties than previously thought. However, milking scenes depicted in prehistoric African rock art and in Saharan ceramics have so far not produced dates or residues that bear on the antiquity of milking in Africa (Jesse et al. 2007; Marshall 2000).
Oscillating periods of aridity and humidity resulted in periods of increased mobility and occasional depopulation of the Sahara (di Lernia 2002; Garcea 2004; Kröpelin et al. 2008). In the eighth to seventh millennia cal BP, herders combined livestock keeping with hunting and collection of wild grain in regions such as the Acacus Mountains (Gautier 1987b). At Adrar Bous and other sites near lowland lakes, herders also fished and collected shellfish (Gifford-Gonzalez 2005; Smith 1992; fig. 1). Cattle-focused rock art attests to the symbolic importance of cattle for Saharan herders (Holl 2004; Smith 1992, 2005). Hunter-gatherers also flourished during this period at sites such as Dakleh Oasis (McDonald 2008) and Amekni (Camps 1969; fig. 1), creating a mosaic of hunters and herders across northeastern Africa (fig. 1).
Through the mid-Holocene, grasslands became more arid, precipitation became increasingly unpredictable, and desert regions of the Sahara expanded. Northeastern Africans responded to these pressures by heightening mobility, relying on introduced sheep and goats, and decreasing use of wild cereals (Barich 2002; di Lernia 2002; Garcea 2004; Gautier 1987a). It was during this period that the donkey was domesticated (Rossel et al. 2008). Their use would have made increased residential mobility and dispersal of settlements from water possible and would have facilitated long-distance migrations (Marshall 2007).
Significant expansion of the geographic distribution of the dotted-wavy-line ceramic motif and distinctive human mortuary practices in the early seventh millennium cal BP reflect the southward movement of pastoralists, long-distance contacts among Saharan groups, and elaboration of pastoralist ideologies (Jesse et al. 2007; Keding, Lenssen-Erz, and Pastoors 2007; Smith 1992; Wendorf and Królik 2001). Just as in the Mediterranean and western Europe, however, the trajectories of small immigrant groups may have varied greatly (Özdoğan 2011; Rowley-Conwy 2011). Domestic stock appear to the south in the Sudanese Sahel by the early seventh millennium cal BP at Esh Shaheinab and Kadero (Gautier 1984a, 1984b) and by the mid-fifth millennium cal BP in Kenya (Marshall and Hildebrand 2002). Similarly, Saharan lithics and other traces of Saharan herders are first found in the West African Sahel by approximately 4500 cal BP (Jousse et al. 2008; Linseele 2010; Smith 1992). Di Lernia (2006) argues that the widespread ritual burial of cattle across the Sahara at the end of the seventh millennium BP represents a social response to rapid aridification. Cattle burials and associated ritual activity are a prominent feature of site E-96-1 at Nabta (Wendorf and Królik 2001). At Djabarona 84/13, in the middle of Wadi Howar from the beginning of the sixth millennium cal BP, more than a thousand pits are filled with cattle bones and relatively complete ceramic pots (Jesse et al. 2007; fig. 1). As far south as Kenya by the middle of the fifth millennium cal BP, large stone circles such as those at Jarigole were constructed as centers for human burial rituals by southward-migrating herders (Marshall, Grillo, and Arco 2011; Nelson 1995). Hunter-gatherers, however, continued to flourish after the movement of herders into these regions (Lane et al. 2007; Lesur, Vigne, and Gutherz 2007).
Domestication of African Plants
The earliest evidence for domestication of indigenous African plants with morphological change dates only to the beginnings of the fourth millennium cal BP (table 1). Although many Holocene hunter-gatherers of northeastern Africa relied heavily on wild Saharan cereals, high mobility and repeated abandonment of the region seem to have impeded long-term directional selection and morphological and genetic change. Instead, selection processes culminated in morphological change once Saharan herders settled in the southern reaches of the Sahara and more humid Sahelian regions and established more permanent settlements in areas that were still within or close to the edge of the wild range of Saharan species.
Sahelian herderswho also hunted, gathered, and fishedintegrated cultivation of domestic pearl millet Pennisetum glaucum into their subsistence economies in one or two domestication events documented at or after 38983640 cal BP at sites west of Lake Chad, including Karkarichinkat Nord (KN05), Dhar Tichitt, Birimi, and Gajiganna (DAndrea, Klee, and Casey 2001; Fuller 2007; Kahlheber and Neumann 2007; Manning et al. 2011; fig. 1, table 1). Morphologically, this is evidenced by changes in seed shedding and shape, although increases in seed size were delayed (DAndrea, Klee, and Casey 2001). Fuller (2007) argues that the appearance of domestic pearl millet in India in the mid-fourth millennium cal BP indicates a somewhat earlier African domestication and rapid dispersal. Recent research has also shown that the cow pea Vigna unguiculata was also an early-fourth-millennium morphological domesticate, dating to ca. 38983475 cal BP at the Kintampo B-sites in the grasslands of central Ghana (DAndrea et al. 2007; table 2). By contrast, African rice Oryza glaberrima was domesticated in the inland Niger delta of the Niger bend region by the early second millennium cal BP. On the eastern side of the continent, domestic teff Eagrostis tef and finger millet Eleusine coracana were cultivated by Aksumite populations in the Ethiopian highlands by the beginnings of the second millennium cal BP (historic date AD 150350; DAndrea 2008). The oil-seed noog Guizotia abyssinica is also present in Late Aksumite contexts (DAndrea 2008). DAndrea (2008) points out, however, that morphological change is difficult to identify in the small-seeded cereal teff, which was selected for reliable production under arid conditions rather than for increased seed size. In humid forested southwestern Ethiopia, Hildebrand (2003a, 2003b, 2007) has documented varied selection processes leading to domestication of yams Dioscorea cayenensis and ensete Ensete ventricosum. In these and other areas of Africa, domestic plants are thought to have been advantageous to pastoral hunter-fishers for risk minimization and greater predictability (DAndrea et al. 2007; Kahlheber and Neumann 2007; Marshall and Hildebrand 2002).
Although morphological change occurred in a range of domesticated African plant taxa, it has been suggested that a number of African savanna plants were cultivated or intensively managed over the long term in ways that did not lead to morphological domestication (reviews in Marshall and Hildebrand 2002; Neumann 2005). Haaland (1999) and Abdel-Magid (1989) argued, largely on the basis of the ∼30,000 grindstones that were unearthed at the site of Um Direiwa, for cultivation of sorghum Sorghum bicolor in Sudanese sites dating to the seventh millennium cal BP (table 1). Mechanisms that they suggested for late morphological change include continued outcrossing between cultivated and wild populations and harvesting through beating into baskets or uprooting. This has led to arguments that sorghum was not morphologically domesticated until it was removed from its wild African range (Haaland 1999; but see Fuller 2003). Although mechanisms exist that may have caused late morphological change in African cereals and harvesting of wild grains was at times intensive, there is no macrobotanical evidence or indication of landscape modification that supports claims for cultivation of African grains before the early fourth millennium cal BP.
In the wetter tropical regions, there is evidence of long-term use of a number of forest taxa without morphological change. Long-term use of oil palm Elaeis guineensis and incense trees Canarium schweinfurthii has been documented across the humid tropics of Africa (DAndrea, Logan, and Watson 2006; Mercader et al. 2006). This pattern is not confined to forests, however. DAndrea, Logan, and Watson (2006:216217) argue that Kintampo people living in the grasslands of central Ghana employed a system of arboriculture that did not rely on management strategies that would result in morphological change. Kahlheber and Neumann (2007) also note that a number of west African park savanna species, such as baobab Adsonia digitata and the shea-butter tree Vitellaria paradoxa, were protected and encouraged but never domesticated. Other wild plants that are still protected and sometimes actively sown in many different African environments include weedy green species ranging in status from crops to semidomesticated or wild (Kahlheber and Neumann 2007; Marshall 2001). Kahlheber and Neumann (2007:333) point out that in the West Africa Sahel, reliance on morphologically wild park savanna species became more evident when economies diversified and populations concentrated close to water 2,000 years ago. In many regions of Africa, Iron Age agriculturalists relied on a particularly broad range of resources, and farmers incorporated diverse domestic crops and managed plants, cattle, sheep, goats, dogs, and donkeys into their agricultural systems and fished and hunted a wide range of wild-animal foods (Casey 2005; Neumann 2005; Plug and Voigt 1985; van Neer 2000).
This brings to the fore the question raised at the outset of whether such diverse subsistence strategies fit current conceptions of agricultural systems. Kahlheber and Neumann (2007:339) are doubtful whether farming is an appropriate term for some of these ways of life. Smiths (2001, 2011) term low-level food production has been used in the region, but it does not fully capture the complexities of African settings. The question of whether the Kintampo should be considered foragers, farmers, or something else has also been reviewed by Casey (2005) and by DAndrea and colleagues (DAndrea, Logan, and Watson 2006:216218; DAndrea et al. 2007), who argue that although there are clear-cut cases of foragers or farmers in Africa, there are many others that defy simple categorization. Hildebrands (2003a) ethnographic research among the Sheko of southwestern Ethiopia and the literature on use of weedy greens in Africa (Etkin 1994; Fleuret 1979; Marshall 2001 and references therein) provide ample evidence that such subsistence strategies have long-term trajectories in many parts of Africa and cannot be dismissed as transitory.
Ethnoarchaeological Insight into Management, Selection Processes, and Domestication of the DonkeyJump To Section...
One approach to better addressing conceptual problems presented by questions of late morphological change and the diversity of economic systems in Africa is to consider pathways to domestication for particular species in light of the potential for morphological change, or lack thereof, in specific social and environmental contexts. The question that we address here is how the behavior of the African wild ass and management of donkeys by herders and small-scale farmers in Africa contribute to selection processes and the likelihood of development of archaeological signatures of domestication in the donkey. This analysis focuses on aspects of the biology and behavior of the donkey and its use as a transport animal that influence management practices in extensive pastoral and agricultural systems and are relevant (sensu Wylie 2002) to ancient settings for domestication. It is often argued, for instance, that sociability and the presence of a dominance hierarchy are desirable characteristics for potential domesticatability (Clutton-Brock 1992; Diamond 1997). African wild ass do not, however, fit this profile. The extant Somali wild ass, or dibokali, is solitary or forms groups with weak short-term associations. It also lacks a pronounced dominance hierarchy (Klingel 1974; Moehlman 2002). This social system profoundly influences donkey behavior under human management.
Recent ethnoarchaeologial research on donkey use and management among Maasai households in Kajiado District of southern Kenya provides the first detailed information on selection processes in a pastoral social and economic context. During 2006, Lior Weissbrod lived in Maasai communities in the study area and collected interview and participant observation data from 26 women from eight households spread among six different pastoral settlements (table 3). The study focused on use and daily management, herd composition, mortality, and breeding behavior. After a 2-year period of severe drought (20042006), the donkey holdings of households participating in the study were reduced but still totaled 65.
Donkeys were not regarded as food. They were considered womens animals, important for transport but without the symbolic status of cattle. Women were the caretakers of donkeys and used them to carry household goods during residential moves, to collect water, and to take intermittent trips to trading centers. Donkeys also carried meat, firewood, and water for large ceremonies. During the dry season, women went long distances for water every other day, returning with a typical load of 50 L per donkey. Children herded household donkeys with the calves, but during the wet season, donkeys were free ranging. Many families penned donkeys within the settlement thorn fence or in calf enclosures at night for protection against predators.
Our data show that the use of donkeys in Kajiado enhanced the flexibility and stability of local herding systems (see also Marshall 2007; Marshall and Weissbrod 2009). Families in the study area who did not own donkeys could not move as a whole away from permanent sources of water and were unable to make optimum use of available grazing. Donkeys were, nevertheless, managed less than other livestock. Marshall (2007) previously noted that the ability of donkeys to dig for water and to protect themselves from predators more successfully than other livestock was associated with low levels of management, which might result in low levels of selection. Our data show that behavior was a factor but that the level of use of donkeys in the study area ultimately determined the degree to which donkeys were herded and penned.
In addition to management practices, we also collected information on reproduction and desired characteristics of donkeys that might be selected for through strategic breeding. Women that we talked to particularly valued strength and calmness in a donkey. Some also mentioned the importance of disease and drought resistance, although they noted that donkeys were less vulnerable to these hazards than other livestock. We found, however, that participants in the study made no attempt at all to influence mate choice among donkeys or to breed for particular characteristics. The ancestry of a particular donkey was unknown except for the female parent. By contrast, research on cattle genealogies shows that Maasai herders memorize these in great detail for several generations (Ryan et al. 2000). The lack of strategic breeding of donkeys is influenced by donkey behavior and herd compositions but is also related, at least in part, to the fact that Maasai herders do not use donkeys as symbols of social transactions in the same way that they do cattle or value color distinctions ideologically.
The dynamics of wild ass mating systems, based on short-lived associations that occur when females move through male territories, influence donkey breeding in the domesticated environment. Maasai women stressed their concern with the aggressive behavior of jacks during mating. Even when they wanted to keep a female from breeding with an especially aggressive jack, women said that they found it impossible to keep the male away. They also noted that estrus jennies might go astray without warning in search of males. They are often lost this way, and we documented a number of cases in which wandering females, as well as males, were cared over a long term by women in distant settlements. Lack of selection because of the difficulty of controlling donkey breeding is, therefore, likely wherever a premium is placed on wild characteristics of the donkey, such as strength, rather than on docility and productivity for food. The relatively high proportion of males in herds (one male∶two females) is another factor that makes control over breeding logistically difficult. Because donkey owners kept small herds specifically for transport, they weighed the breeding advantages of females against the superior transport potential of males. The strength of males was greatly favored, and so was their consistent availability for transport use.
Herd growth and mortality patterns also contributed to patterns of selection in domestic donkey herds. Pastoral Maasai donkeys had, on average, a foal every 2 years. Mortality resulted from predation by hyenas, disease, and drought. Herds grew relatively slowly, and additional animals were recruited to herds through gifts, loans, and purchase. Socially based loans or exchanges of cattle are deeply woven into the fabric of Maasai society (Ryan et al. 2000). To a lesser extent, this system is also used for donkeys, and social exchange is a mechanism of selection and gene flow. Animals entering or leaving a herd through loans were carefully selected and predominantly female. In some cases, however, exchanges were involuntary, resulting from donkeys running away.
In the wet season, herds of donkeys made up of animals from different settlements in the same neighborhoods range freely. This practice and the system of intentional and unintentional loans maintain gene flow among settlements. Purchases were rarer than loans also but recruited animals to slow-growing herds and maintained intentional selection on an interregional scale. Men purchased animals when visiting markets, and the strength and price of the donkey were major considerations affecting purchases. Young male donkeys were cheaper than others, and purchases were one male∶two females. There was no intentional culling of donkeys, and donkeys were not eaten, but small, slow-growing, or aggressive males were removed from the breeding pool through castration. We recorded six castrated males (40% of the males studied), and castration of male donkeys was a more important factor affecting the direction of intentional selection than culling or selective breeding.
Very few studies of donkey management and selection have been conducted in settled agricultural villages. Mohammeds (1991) and Wilsons (1991) Ethiopian research can, however, be used for comparison with the Maasai pastoral study. They focused on Ethiopian farmers of the central and southern highlands who used donkeys to transport grain to market and for hauling household firewood and water. Most families in the study areas kept one to two donkeys, usually female (Mohammed 1991; Wilson 1991). Donkeys were also loaned to family and friends. In the Awassa region, males were rare (100 females∶1 male); in other regions the number of males was higher (73 females∶27 males). Where males were more common, they were usually less than 4 years old. Mohammed (1991) notes that male donkeys were not castrated. We infer that low proportions of males in herds indicated male culling, although donkey eating was not discussed. People in Awassa did not supervise donkeys when they were not using them, and Mohammed (1991) documents minimal donkey management and poor animal nutrition in this area. Because of the danger presented by hyenas, however, people often brought donkeys inside their houses at night. There was no intentional control over breeding, however. Mohammed mentions that copulation might occur anywhere and was actively discouraged in the market center (Mohammed 1991).
Overview of management and selection.
In order to consider patterns of directional selection, it is useful to examine factors that affect the likelihood of genetic drift, intentional selection, and reproductive isolation in donkeys managed by pastoralists and small-scale farmers. Culling of male donkeys by Ethiopian villagers and castration of male donkeys by Maasai pastoralists were important factors affecting selection. These practices ensured that males with desired traits, such as strength or size, remained in the breeding pool. Females, on the other hand, were never culled, and management of donkeys was minimal. None of the donkey owners that we studied tried to ensure a diverse set of breeding males, to breed select females or males, or to keep records of parentage. We argue that these management practices are influenced by wild ass and donkey courtship and breeding behavior and have significant consequences for long-term directional selection and domesticatory processes. The data also indicate that different sets of functional and symbolic considerations affect Maasai practices of cattle and donkey management and are associated with differing levels of selective pressure and control of gene flow. In our study area, people also bred or obtained cattle for ideal coat colors and conformation, and it is possible that without this additional symbolic motivation, functional reasons for breeding donkeys were not enough to overcome significant practical difficulties. As research on mammals such as the fur fox (Belyaev 1979; Trut 1999) and the guinea pig (Künzl et al. 2003) has shown, without selective breeding, retention of individual animals with desired traits and culling of others, directional selection may be very slow or fail to occur even in the absence of gene flow from wild populations.
From a wider perspective, there are related issues that work against genetic drift as a major factor driving genetic and morphological change in donkeys. In both the Maasai and Ethiopian Arsi cases, donkeys from numerous households grazed unsupervised in mixed herds, allowing uncontrolled genetic exchange among neighborhood populations. Donkeys were loaned among broad social networks in both regions, and the frequency with which donkeys were taken to market in Ethiopia also provided a wider setting for interbreeding among donkeys from different areas. We argue, however, that in both the pastoral Maasai and Arsi farmer cases, low levels of formal management and lack of intentional selective breeding are linked to donkey biology and behavior, the use of donkeys for transport, and the fact that donkeys are not often eaten. Male culling plays a significantly greater role in animals that are primarily managed for meatincluding cattle, sheep, and goatthan it does in donkeys. Although culling and castration affect donkey selection, they are outweighed by lack of directional selection in breeding and consistent gene flow among donkeys over significant distances.
The data for Maasai pastoralists and Ethiopian Arsi farmers also suggest that the potential for gene flow from the wild is likely in both settings but marginally less so in agricultural villages. The Maasai villages studied lie outside the historic range of the wild ass. But it is easy to see that had they not, the runaway tendencies of estrus females would have made the prevention of introgression difficult. Like contemporary herders valuing strength and endurance in their donkeys, historic Beja pastoralists of Sudan and Eritrea intentionally encouraged interbreeding among donkeys from domesticated and wild settings (Baker 1867; Murray 1935). During the 1950s, Nicolaisen (1963) also recorded capture and taming of wild or feral animals by Tuareg pastoralists of the central Sahara.
It is possible, therefore, to begin to identify separate contexts for the domestication process of donkeys in Africa. We predict that ancient Saharan pastoralists reduced the number of breeding males in herds through culling and castration in order to cope with practical difficulties resulting from courtship and breeding behavior in donkeys. Isolation from wild ancestors would have been possible in some pastoral settings as a result of mid-to-Late Holocene climate change, range fragmentation, and pastoral settlement in island or marginal ecosystems. Wild asses may also have been removed from their wild range by pastoral dispersals into the high-altitude Ethiopian highlands and other regions, such as southern Sudan and northern Kenya, outside the historic range of the wild ass.
Selection for morphological change would have been slow until donkeys were removed from close proximity to the wild ass and interbreeding between local donkey populations was restricted. It would appear that reproductive isolation of captive wild asses from free-living populations is somewhat more likely to have occurred in ancient urban settings such as the Predynastic and Dynastic Egyptian towns of the Nile Valley, with permanent walls and high densities of protected agricultural land. Gene flow would still have been possible, however, given the narrowness of the Nile agricultural belt and the mobility of pack donkeys. An appreciation for the advantages of strong animals may also have made interbreeding between captive and wild asses desirable for both villagers and pastoralists.
The lack of morphological change evident in the Abydos donkeys as late as 5000 cal BP (3000 BC; Rossel et al. 2008) demonstrates that size decrease was not generally established until well after this period. It is also conceivable that morphological change did not occur until donkeys were taken across the Red Sea to Yemen or other regions of Asia. Whichever the case, donkeys are a classic example of a species that was used to carry loads for millennia as a domesticate but with late morphological change. We conclude that slow morphological change in domesticated donkeys can be explained by low levels of selection, high potential for interbreeding between founder populations, and potential for introgression with the wild.
Do Holocene Pastoralists in Africa Fit Conceptions of Early Agricultural Systems in Other Regions?
After examining evidence for the beginnings and spread of food production in Africa and analysis of the way that management and behavioral factors affect the likelihood of morphological change in one large mammalthe donkeywe return to consideration of whether African pastoralism fits current conceptions of early agricultural systems developed for other regions. We start by considering the question of whether recognition of early food production in tropical regions of Africa has been hampered by concepts of domestication that rely on morphological change by focusing on donkeys, cattle, Barbary sheep, African cereals, and West African tropical tree crops.
Some evidence suggests that complex hunter-gathers may have attempted to manage cattle in the northeastern Sahara and, for a time, Barbary sheep in the Libyan Acacus. There is no doubt that short-term participation in domesticatory relations are difficult to recognize archaeologically, but nevertheless evidence for management of Barbary sheep is suggestive rather than conclusive. In contrast, genetic data offer a measure of support for the hypothesis of cattle domestication in Africa. The sociality of wild Bovini, however, and the expectation that wild cattle were used mainly for food suggests strong selection and a pathway to domesticationcharacterized by a postmanagement lag rather than late morphological change and fewer problems with identification of early domesticatesdifferent from that discussed for the donkey.
Ethnoarchaeological data on the donkey reveal relations among selection processes and slow genetic and morphological change and illuminate conditions under which biology and human management influenced domestication and the likelihood of late morphological change. The biological and behavioral reality of donkeys in current domesticatory settings in Africa is that females actively seek out mates, territorial males are reproductively aggressive, and high proportions of males are advantageous for transport use. These factors interact to make reproduction difficult to control and gene flow likely among donkeys of different households and villages, along trade routes, and between tame animals and wild asses.
Archaeological and genetic data suggest that pastoral societies of the Sahara or the Horn of Africa played an important role in the early development of stable and long-term systems of management of morphologically wild donkeys. Morphological change was late, and mechanisms for this probably included creation of built environments of the Nile Valley, late agriculturally modified landscapes, the high mobility of Saharan pastoralists, and ecological fragmentation created by climatic changes of the mid-Holocene.
Although an appreciation of the likelihood of delayed morphological change and biases against identification of domestic donkeys is novel, Africanists have long discussed the question of whether the lack of morphological change resulted in bias against recognition of cultivation of early cereal crops. There is mounting evidence for long periods of intensive use of wild cereal grasses by Early Holocene hunter-gatherers and early herders of the Sahara without evidence of domestic traits. This has been related to a lack of continuous directional selection as a result of increased aridity and pastoral mobility. Morphological changes in well-known African cereals such as pearl millet and pulses such as cowpeas occur relatively late and in conjunction with pastoral sedentization in better-watered locales within the semiarid Sahel and in the more humid West African woodlands after the fourth millennium cal BP. Recent research in more humid regions of West Africa has revealed, however, a number of tended and managed tree crops, such as incense, baobab, and the shea-butter tree, that were heavily used during the Holocene but remain morphologically wild to this day. This is typical of tropical tree crops worldwide and common in weedy greens.
It is worth reiterating at this point that identification of management of plants and animals before genetic or morphological change is inherently problematic, and the longer the period before morphological change occurs in a particular plant, animal, or setting, the greater the difficulties that arise. It is clear that there are at least three axes of variability in morphological responses of plants and animals to selection during coevolutionary relations with humans. We have found it useful here to conceive of this temporal and spatial variation in terms of a postmanagement lag before morphological change, as opposed to late morphological change or regionally clustered variability.
Our review suggests that all these forms of variability exist in Africa. The available data appear to accord with Jones and Browns (2007) suggestion that a long, stable period of management without morphological change or a normal morphological lag is common to many domesticates worldwide. In Africa, however, it is not clear that their corollarythat population expansion leads to removal of plants and animals from their wild range and morphological changeholds true. Instead, heightened mobility related to climatic changes and increased aridity ultimately led to the movement of some species out of their wild ranges. Furthermore, early African cereals appear to have been domesticated within their wild ranges and intensified on the edge of these regions. Increasingly settled pastoral communities and management practices that maintained directional selection seem to have been more important factors affecting domestication of these crops than reproductive isolation.
We focused above on the possibility of biases against the recognition of early agriculture in tropical regions. We do not, however, see a cluster of taxa subject to late morphological change in the arid or high-altitude subtropics of Africa; here, species-specific analyses of the likelihood of late morphological change are crucial. We agree with Denham (2007), however, that the biology of many species of the African humid tropics increases the likelihood of a lack or significant delay of morphological change and the potential for interpretive bias. These data are strongest with regard to African tree crops. Despite this, however, there is little evidence that archaeologists have ignored early agriculture in the humid tropics of Africa. There is, in fact, no archaeological evidence that the humid tropical forests were heavily populated by African hunter-gatherers during the Early Holocene, and there are few traces of intensification in these regions until after they were settled by food producers (see DAndrea, Logan, and Watson 2006; DAndrea et al. 2007; Mercader et al. 2006 and references therein). Nevertheless, as Africanist paleoethnobotanists have pointed out, much work remains to be done on the nature of agricultural systems dating to the past several thousand years in the humid tropics (DAndrea, Logan, and Watson 2006; Hildebrand 2007; Kahlheber and Neumann 2007).
We conclude that there is no indication of significant regional-scale biases that would have affected current interpretations of the sequence of plant and animal domestication in Africa or geographic patterns of the timing and spread of food production. The larger patterns, as we see them, are that some complex hunter-gatherers of the Early Holocene in North Africa successfully managed cattle, developed pastoral social and subsistence systems, and spread over vast areas of the Sahara. Other such groups in North Africa may have experimented with management of Barbary sheep, but this was short-lived. Later, during the mid-Holocene, there is evidence that donkeys were domesticated by African pastoralists in the Sahara and the Horn of Africa and possibly by Predynastic Egyptians in towns along the Nile. These animals remained morphologically wild for long periods. The earliest plant domesticates in Africa are associated with decreased mobility as pastoralists moved into better-watered locales within the semiarid Sahel and into West Africa. It can also be shown, however, that in some humid tropical regions of Africa, clusters of species existed with a long history of cultivation or tending by established agricultural communities and with biological traits amenable to management but no traces of morphological domestication.
African patterns of food production were distinctive. Animals were domesticated before plants, herding populations became more mobile than their forager ancestors, the subsistence system was characterized by a few morphologically wild domesticates (e.g., the donkey), a wide range of wild resources in ecodiverse combinations continued in use, and mosaics of hunter-gatherers and herders occupied varied regions. Pastoralism developed early in the arid topics, whereas the beginning of farming based on domesticated plants was late.
These African data are informed by and provide perspectives on pathways to food production in other regions. In discussions at the Wenner-Gren conference in Temozón in 2009, Meadow (2009) and Fuller (2009; also see Fuller 2006) argued that South Indian patterns of early pastoralism and subsequent domestication of local millets and pulses are reminiscent of Africa. Similarly, pastoralism has long been considered an early phenomenon in the Andes (Aldenderfer 2003; Browman 1974; Mengoni-Goñalons and Yacobaccio 2006) and the Zagros (Abdi 2003; Hole 1996). Mobile pastoralism is also a major theme in data emerging on the beginnings of food production in central Asia (Frachetti and Benecke 2009; Outram et al. 2009). In addition, Belfer-Cohen and Goring-Morris (2011) and Goring-Morris and Belfer-Cohen (2011) document African-like mosaics of hunter-gatherer and early-food-producer settlement in the Levant during the Early Holocene. Evidence is also mounting that shows continued reliance on wild resources and ecodiverse strategies pursued by small-scale food producers or low-level farmers of the Americas and subtropical and tropical regions (Denham 2011; Fritz 2007; Piperno and Pearsall 1998; Smith 2001, 2011) and perhaps even temperate regions of Asia (Crawford 2011; Lee 2011; Zhao 2011). Understanding ways in which specific strands such as these contribute to larger similarities and differences in the warp and weft of data on the beginnings of agriculture requires attention to methods of detection of early phases of domestication, information on specific social contexts, and regionally focused and temporarily expansive research. These kinds of data are only just beginning to emerge from Africa, which, as this summary demonstrates, has much to contribute to unraveling patterns of variability in global pathways to food production.
Acknowledgments
We are grateful to Ofer Bar-Yosef, Douglas Price, the Wenner-Gren Foundation, and our companions in Temozón for the genesis of this paper and for challenging and enriching our views about domestication and the spread of food production. The donkey ethnoarchaeological research could not have been conducted without the expert knowledge and gracious support of the people of Kajiado. We are also grateful to the Kenya National Museums and Dr. Purity Kiura for their support and to the government of Kenya for permission to undertake research. This paper has benefited greatly from the thoughtful comments of reviewers. Research was supported by National Science Foundation grants BCS-0447369 and BCS-0536507. [/QB]
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
posted
^^^ inability to use his brian to critique the article that is topic of thread, resorts to hiding behind copy and paste to try to prove he's smart. Other posters formulate arguments in their own words and use their own analysis in reply to a given article instead of just posting different copied info, same info over and over again read it before. If he couldn't rely on this spam he would be lost. I hate name droppers This idot follows me around like a puppy dog
A brand new 2013 article on Africa that Asante Korton also refernced and this fool has got to piss on every territory he sees like a dumb dog or kid that carves his name in a school desk
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
posted
^LOL my intellect exceeds yours by far. LOL
Merely a few posts destroyed your Eurocentric nonsense about MY PEOPLE.
Even your own post disputes you. And you should be glade that I posted the sources I did. Since they elaborate on this issue.
quote: This study provides the first strong quantitative evidence for the presence of three major river systems flowing across the Sahara during MIS 5e.
--Jorge A. Ramirez
quote:Originally posted by Djehuti: Here is a paper that was cited years ago in this forum:
When the Sahara Turned from Green to Brown: Post-glacial Climate Change and Human Settlement in Central Sahara, 12,000 - 2,500 BC. Abstract: For decades, archeologists have been aware of evidence suggesting that the Sahara Desert was much wetter and greener thousands of years ago. It is now possible to characterize these locations, in terms of both aquatic and terrestrial biota, but by also providing some data on the human inhabitants of these ecotomes. This thesis focused primarily on the features of human settlement in the central Sahara, looking at lifestyles of the people and examining factors favoring a wetlands economy. Finally, as the once-favorable conditions began disappearing, analysis of the destiny of these Saharan peoples is made.
The Holocene followed the last glacial age, about 12,000 BC. The temperatures in the Sahara became appreciably warmer and the climate demonstrated a lower evapotranspiration rate. The Intertropical Convergence Zone (ITCZ), the line along which north and south weather fronts converge, seems to have moved several hundred miles to the north. Since, in general, African rainfall north of the equator increases as one heads south, the shift of the ICTZ resulted in Saharan and sahelian zones receiving increased rainfall.
In addition to more temperate plant and animal species moving into the Sahara, the increased moisture resulted in lakes and rivers filling far beyond their prior capacity. In some cases, rivers breached their normal watershed, connecting with other systems. This allowed a broader distribution of aquatic species. In some areas, there were widespread wetlands. The earliest such sites, when associated with human habitation, date from ~7000 BC (with harpoons) and 6000 BC (wavy-line pottery). Because they used pottery, these cultures were originally considered some kind of Aquatic Neolithic.
JEG Sutton, in a 1974 article, called them the "Aquatic Civilization of Middle Africa". Although they did little stonework, he saw their bone work as very sophisticated. He suspected that much of their material culture, made of perishable materials, would not have survived. He thought them "victims" of stone-oriented scholars. Based on geographic distribution. he thought that the ancestors of Saharan, Songhai and Chari-Nile-speakers were these same "Middle Africans".
After the initial Holocene (postglacial) wet phase (12,000-7000 BC), conditions became drier for 500-1000 years. This was fol- lowed by a lesser wet phase of 2000 years. From then until 2500 BC, there was a gradual increase in dryness. After that time, weather conditions have been largely unchanged until the present. How did the human economies change with the climate?
The early harpoon-fishers availed themselves of large riverine and lake species, such as the Nile perch (known to exceed six feet). Whether they were fully-settled or did some amount of yearly travel isn't known. Undoubtedly, as water-tables dropped, the people needed to augment their aquatic diet with other foods. Initially, these would be available plant and animal foods which could be gathered.
Christopher Ehret's work touched on Sutton's language hypothesis--that these early fishers were NiloSaharan speakers. In the process of generating protolanguages for these speakers, he created an initial vocabulary for both NiloSaharan, proto-Saharan and proto-Sahelian. The words that he developed do not include fish or fishing terms (line, net, hook, harpoon). This early vocabulary does have herding terms and words for 'goat', 'young goat', 'cow', 'corral' and such. No other writers dealt with this topic.
The Central Sahara is punctuated by large stone outcrops--massifs--that create their own microclimate. In some cases, they represent a refuge for rare species, extinct everywhere else. Work by Henri Lhote in the 1950's documented the varied and colorful rockpaintings found in the massifs. Grouped into 23 styles, they are grouped as 1. Bubalus (extinct buffalo) hunters, early Neo- lithic; 2. 'Bovidian' pastoralists (considered Neolithic); 3. Equine phase, involving pastoralists with chariots and cavalry; 4. Phase of the camel, 1 AD. These groupings are disputed, but the images show us a wild-animal phase, then so-called 'Roundhead' figures. These appear to be multi-ethnic, including a 'Negroid' population.
The pastoralist period involved humpless cattle with large curved horns, similar to the ancient Egyptian Longhorn. Features of the people's life resemble activities of the Peul/Fulani, a contemporary West African pastoral group. Some of the boat images re- semble those on Egyptian monuments, raising yet other possibilities. Pastoralism appeared about the time that weather condi- tions became considerably drier (7000 BC), suggesting that it was a selected response to changing conditions.
The equine phase and its suspected timing may represent the arrival of the "Peoples of the Sea", groups that left Crete about 1500 BC (around the time of the Trojan War) and migrated northeast, east and southeast. The Biblical Philistines are one such group. Cretans arrived in NE Africa about 1200 BC, joining with the Libyans and attacking Egypt. Similar groups may have moved in from the coast and entered the central Sahara via the caravan trails. If so, this movement is more a political one than a response to climate change in the central Sahara, although its outcome would affect the local population.
Although the rock art isn't datable, it nevertheless shows us several different worlds of the central Sahara. The earliest involves Africa's megafauna (lions, elephants, etc) and Negroid peoples. Another one shows herding people with cattle. Some features of these images recall West Africa, others, Egypt. The herders look more like Somali people, with narrow noses, thin lips, straight hair. Probable intruders with horses and chariots are seen and, finally, the camel makes its appearance. Are the earlier people migrants from North Africa, from Egypt or from elsewhere?
Dhar Tichitt in southern Mauritania has been instructive as showing the cultural response to a drier climate. Digging revealed eight phases, from hunting megafauna (2000 BC), to limited hunting, gathering and herding (1500-1100). Subsequent phases included significant milling. The involved plant went from cramcram, a spiny famine food, to millet and sorghum. Identification of the species showed that the people had switched from gathering wild grasses to planting them, in about 100 years. Such speed is unheard of under normal circumstances, and suggests that the people were somehow "presensitized" to cultivation, perhaps via a smaller outgroup that grew up with farming and then migrated here. Both herding and planting were presumably responses to unfavorable climate. The site was abandoned after horses and metal weapons arrived, possibly with the charioteers described above.
The future of the central Saharans was not always the same as at Dhar Tichitt. Evidence suggests that the people migrated, some southwest, some southeast, some perhaps north, following the drying riverbeds as they sought sites where they could sus- tain themselves. Since West Africa had not yet been favorable to settlement, due to its dense forests, the central Saharans may represent some of the early ancestors of some of these peoples.
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
posted
A map of what the Sahara was like 8,000 to 11,000 years ago, revealing rivers, lakes, swamps and fan-shaped deposits where rivers emptied out. The dots indicate where various kinds of pottery were found.
Ancient people moving northwards out of Africa had more routes open to them than previously thought, according to research published today.
A generalised map of the Sahara shows the location of the sample sites and the fossilised river courses.
A photograph from NASA's space shuttle Endeavour of the Libyan desert showing ancient rivers (1994).
Late Pleistocene and Holocene drought events at Lake Tana, the source of the Blue Nile
Michael H. Marshall et al.
Abstract Magnetic and geochemical core data spanning the last 17,000 years are correlated with new seismic stratigraphy from Lake Tana, Ethiopia, to infer past lake-level change and hence effective precipitation. The data confirm that low lake-level coincides with Heinrich Event 1 (H1) in the North Atlantic, as previously shown from diatom and pollen evidence (Lamb et al., 2007). The lake deepened at 15.3 cal kyr BP and abruptly returned to freshwater conditions, when the lake overflowed into the Blue Nile. Low runoff and lake levels and therefore rainfall are inferred between 13.0 and 12.5 cal kyr BP and may represent southerly suppression of the ITCZ and the associated monsoon front at the time of the Younger Dryas. Two drought episodes occurred at 8.4 and 7.5 cal kyr BP, and are also interpreted as a southward shift in the monsoon front. The first of these events appears to have preceded and been more significant than the 8.2 cal kyr BP. Precipitation declined after 6.8 cal kyr BP, although we do not see an abrupt end to the African Humid Period. This period culminated in a dry episode at ~ 4.2 cal kyr BP, supporting the view that reduced Nile flow was a contributing factor to the demise of the Egyptian Old Kingdom.
Highlights ► 17,000 years of climate change at the source of the Blue Nile, Lake Tana, Ethiopia. ► Southerly suppression of the ITCZ and monsoon front at the time of the Younger Dryas. ► Drought at 8.4 cal kyr BP preceded and was more significant than the 8.2 event. ► No abrupt end to the so-called African Humid Period. ► Drought at Nile source a factor in the demise of the Egyptian Old Kingdom.
posted
Science. 2006 Aug 11;313(5788):803-7. Epub 2006 Jul 20.
Climate-controlled Holocene occupation in the Sahara: motor of Africa's evolution.
Kuper R, Kröpelin S.
Source
Collaborative Research Center 389 (ACACIA), University of Cologne, Institute of Prehistoric Archaeology, Africa Research Unit, Jennerstrasse 8, 50823 Köln, Germany.
Abstract
Radiocarbon data from 150 archaeological excavations in the now hyper-arid Eastern Sahara of Egypt, Sudan, Libya, and Chad reveal close links between climatic variations and prehistoric occupation during the past 12,000 years. Synoptic multiple-indicator views for major time slices demonstrate the transition from initial settlement after the sudden onset of humid conditions at 8500 B.C.E. to the exodus resulting from gradual desiccation since 5300 B.C.E. Southward shifting of the desert margin helped trigger the emergence of pharaonic civilization along the Nile, influenced the spread of pastoralism throughout the continent, and affects sub-Saharan Africa to the present day.
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
posted
Am J Phys Anthropol. 2011 Sep;146(1):49-61. doi: 10.1002/ajpa.21542.
Biogeochemical inferences of mobility of early Holocene fisher-foragers from the Southern Sahara Desert.
Stojanowski CM, Knudson KJ.
Source
Center for Bioarchaeological Research, School of Human Evolution and Social Change, Arizona State University, Tempe, AZ 85287, USA. christopher.stojanowski@asu.edu
Abstract
North Africa is increasingly seen as an important context for understanding modern human evolution and reconstructing biocultural adaptations. The Sahara, in particular, witnessed a fluorescence of hunter-gatherer settlement at the onset of the Holocene after an extended occupational hiatus. Subsequent subsistence changes through the Holocene are contrary to those documented in other areas where mobile foraging gave way to settled agricultural village life. In North Africa, extractive fishing and hunting was supplanted by cattle and caprine pastoralism under deteriorating climatic conditions. Therefore, the initial stage of food production in North Africa witnessed a likely increase in mobility. However, there are few studies of paleomobility in Early Holocene hunter-gatherer Saharan populations and the degree of mobility is generally assumed. Here, we present radiogenic strontium isotope ratios from Early Holocene fisher-forager peoples from the site of Gobero, central Niger, southern Sahara Desert. Data indicate a relatively homogeneous radiogenic strontium isotope signature for this hunter-gather population with limited variability exhibited throughout the life course or among different individuals. Although the overall signature was local, some variation in the radiogenic strontium isotope data likely reflects transhumance into the nearby Aïr Massif. Data from Gobero were significantly less variable than in other worldwide hunter-gatherer populations, including those thought to be fairly sedentary. Strontium data from Gobero were also significantly different from contemporaneous sites in southwestern Libya. These patterns are discussed with respect to archaeological models of community organization and technological evolution.
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
John E. Yellen National Science Foundation, 4201 Wilson Boulevard, Arlington, Virginia 22230
Abstract Examination of African barbed bone points recovered from Holocene sites provides a context to interpret three Late Pleistocene occurrences from Katanda and Ishango, Zaire, and White Paintings Shelter, Botswana. In sites dated to ca. 10,000 BP and younger, such artifacts are found widely distributed across the Sahara Desert, the Sahel, the Nile, and the East African Lakes. They are present in both ceramic and aceramic contexts, sometimes associated with domesticates. The almost-universal presence of fish remains indicates a subsistence adaptation which incorporates a riverine/lacustrine component. Typologically these points exhibit sufficient similarity in form and method of manufacture to be subsumed within a single African tradition.They are absent at Fayum, where a distinct Natufian form occurs. Specimens dating to ca. 20,000 BP at Ishango, possibly a similar age at White Paintings Shelter, and up to 90,000 BP at Katanda clearly fall within this same African tradition and thus indicate a very long-term continuity which crosses traditionally conceived sub-Saharan cultural boundaries.
And more recent sources:
Volume 300, 25 June 2013, Pages 153170
The Middle Palaeolithic in the Desert
The Middle Stone Age of the Central Sahara: Biogeographical opportunities and technological strategies in later human evolution
quote: The makers of these assemblages can therefore be seen as (1) a group of Homo sapiens predating and/or contemporary to the out-of-Africa exodus of the species, and (2) geographically one of the (if not the) closest from the main gate to Eurasia at the northeastern corner of the African continent.
Although Moroccan specimens have been discovered far away from this area, they may provide us with one of the best proxies of the African groups that expanded into Eurasia[...]
quote: The area differs from other sties areas such as the Nile Valley or the Near East because the Middle/Late Paleolithic transition in the Sahara is not marked by changes in core technology. The overall dates for the Libya sites containing the Aterian tool technique range from 47,000- 24,500 BP. Some of the dating techniques were Thermoluminescence (TL) which proved successful in dating several types of sediments including "desert loss" sand dunes.






 UBBFriend: Email this page to someone!
UBBFriend: Email this page to someone!






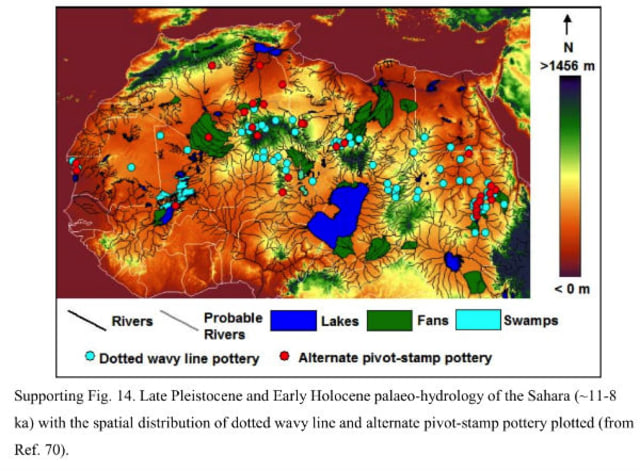














 Printer-friendly view of this topic
Printer-friendly view of this topic