Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans
Maanasa Raghavan, et al
Nature 505, 8791 (02 January 2014) doi:10.1038/nature12736 Received 14 July 2013 Accepted 04 October 2013 Published online 20 November 2013
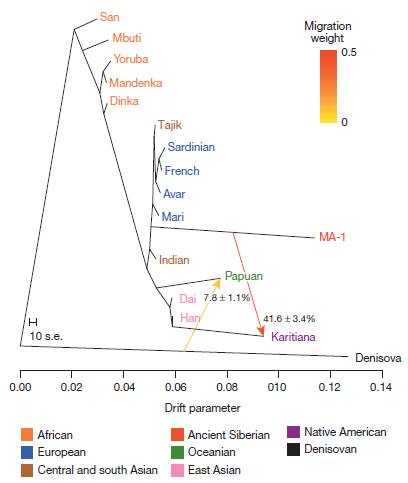
The origins of the First Americans remain contentious. Although Native Americans seem to be genetically most closely related to east Asians1, 2, 3, there is no consensus with regard to which specific Old World populations they are closest to4, 5, 6, 7, 8. Here we sequence the draft genome of an approximately 24,000-year-old individual (MA-1), from Malta in south-central Siberia9, to an average depth of 1×. To our knowledge this is the oldest anatomically modern human genome reported to date. The MA-1 mitochondrial genome belongs to haplogroup U, which has also been found at high frequency among Upper Palaeolithic and Mesolithic European hunter-gatherers10, 11, 12, and the Y chromosome of MA-1 is basal to modern-day western Eurasians and near the root of most Native American lineages5. Similarly, we find autosomal evidence that MA-1 is basal to modern-day western Eurasians and genetically closely related to modern-day Native Americans, with no close affinity to east Asians. This suggests that populations related to contemporary western Eurasians had a more north-easterly distribution 24,000 years ago than commonly thought. Furthermore, we estimate that 14 to 38% of Native American ancestry may originate through gene flow from this ancient population. This is likely to have occurred after the divergence of Native American ancestors from east Asian ancestors, but before the diversification of Native American populations in the New World. Gene flow from the MA-1 lineage into Native American ancestors could explain why several crania from the First Americans have been reported as bearing morphological characteristics that do not resemble those of east Asians2, 13. Sequencing of another south-central Siberian, Afontova Gora-2 dating to approximately 17,000 years ago14, revealed similar autosomal genetic signatures as MA-1, suggesting that the region was continuously occupied by humans throughout the Last Glacial Maximum. Our findings reveal that western Eurasian genetic signatures in modern-day Native Americans derive not only from post-Columbian admixture, as commonly thought, but also from a mixed ancestry of the First Americans. Figure 2 | Admixture graph for MA-1 and 16 complete genomes. An admixture graph with two migration edges (depicted by arrows) was fitted using TreeMix21 to relate MA-1 to 11 modern genomes from worldwide populations22, 4 modern genomes produced in this study (Avar, Mari, Indian and Tajik), and the Denisova genome22. Trees without migration, graphs with different number of migration edges, and residual matrices are shown in Supplementary Information, section 11. The drift parameter is proportional to 2Ne generations, whereNe is the effective population size. The migration weight represents the fraction of ancestry derived from the migration edge. The scale bar shows ten times the average standard error (s.e.) of the entries in the sample covariance matrix. Note that the length of the branch leading toMA-1 is affected by this ancient genome being represented by haploid genotypes.
posted
I'm not at all surprised by these findings. Even in Siberia today there is a present frequency of NRY R* and derived haplogroups. I remember a study showing that Neolithic to Bronze Age Mongolians especially around the Altai area carried R*. Also, it is pretty much established FACT that the Americas were populated by waves of different populations. Is anyone aware that even mt DNA X haplogroups are found in some Native Americans as well?
Posts: 26238 | From: Atlanta, Georgia, USA | Registered: Feb 2005
| IP: Logged |
posted
I am not sure what point you are making here Lioness. But NRY R* .and Q is typical East Asian. Infact if you looked at the data and read the ENTIRE report and Supp you would understand that.
Maybe the experts on here can compare the hg-U haplotypes of MA-1 to modern populations. I am especially interested in the Fulani dataset published by Perrieria et al ( that Berber connection Saami paper). If I had the time there is so much I would do with these dataset. Check out the Supp, page 42 on MA-1 for the SNPs. Man if I had the tools and time!
The Fulani hg-U were also shown to be basal.
Anywho - Here some exceprts fro MA-1 :
================
SI 7. mtDNA haplogroup of MA-1
carries all the three SNPs characterizing hg U (A11467G, A12308G and G12372A). For comparison, we included in the analysis the individual Dolni Vestonice 14 (DV- 14; GenBank accession number KC521458), which has been shown to be basal to the extant hg U51. Both the MA-1 and DV-14 sequences were analyzed for the presence of diagnostic mutations of the major sub-hgs of extant hg U lineages, using information from mtDNA tree Build 15 (Sept 30, 2012)2. A phylogenetic tree was built, with the age estimates (kiloyears, +/- SD) of different sub-hgs of hg U3, including all major extant branches of mtDNA hg U lineages from its root and also the two ancient hg U lineages (Figure SI 4a). The mtDNA sequences of MA-1 and DV-14 share only the three basal mutations inside hg U with each other, and do not belong to any known modern branch of hg U.
To show the present spread of hg U and its different sub-hgs (Figure SI 4b), the average frequencies, divided into four frequency classes, were calculated in regional groups, using a dataset consisting of ca. 30,000 partial mtDNA genomes4-78. Today, hg U is a pan western Eurasian haplogroup, with distribution across Europe, the Middle East, South and Central Asia, western Siberia and North Africa (Figure SI 4b). The overall frequency of hg U is low or absent in extant central and south Siberian populations, i.e. the region close to where MA-1 originated, as well as in East Asia (Figure SI 4b).
SI 8. Y chromosome haplogroup of MA-1 ! Due to low depth-of-coverage of the MA-1 individual (1.5X on 5.8 million bases), genotyping at each site on the Y chromosome was performed by selecting the allele with the highest frequency of bases with a base quality of 13 or higher.
. MA-1 Y chromosome data was then included, and MEGA phylogenetic software3 was used to construct a Neighbor Joining (NJ) tree with default parameters (Figure SI 5a). MA-1 is placed as a basal lineage to hg R2,4. Phylogenetically informative positions and their state in MA-1 were then determined to confirm the placement of MA-1 on the tree.
This resulted in 4301 positions being retained that were classified according to their hg affiliations. Among those phylogenetically informative positions, 1889 non-N positions were retrieved from MA-1. When counting from the split of hg DE on the unrooted phylogenetic tree, MA-1 is determined to be carrying the derived allele in 183 sites and the ancestral allele in 1706 sites. The position of MA-1 on the phylogenetic tree is established by the state of the 313 basal mutations separating hgs DE and R, where MA-1 has 143 informative positions. Of these, 138 are in the derived and 5 in the ancestral state, placing MA-1 as a lineage ***BASAL*** to hg R. With only a FEW EXCEPTIONS characterized below, all other informative positions in MA-1 are in the ancestral state, further supporting the phylogenetic positioning of MA-1 on the tree. Among the derived markers in the final dataset only a few (11) mutations were detected that are likely to be false positives based on the phylogenetic analysis, where it is ASSUMED that recurrent mutation is less likely than a sequencing error. One position among the 35 private to MA-1 is characteristic of a distant hg namely C3c14. Based on current data, 10 additional phylogenetically non-concordant positions in MA-1 were found 1 position for hgs E, G, Q, R1b, R1 each, 2 defining positions for hg I and 3 private mutations for R1b individuals (shown in red on Figure SI 5a). Additionally, among the mutations originally excluded (the reference-private mutations), two positions were found where MA-1 is in derived state.
------
MA-1 could be R* or Q. Typical East Asian
It is becoming very clear these EuroCentric geneticist play games. Eg when they want to steal African history they label the AEians Caucasoid. When they want to steal African genes they label it non-African then use the high frequency in Europe to support their thievery. In the above example they use the high frequency of hg-U to steal MA-1 ignoring the age and the haplotype diversity and relatedness. Also U5 has high frequency in the Scandinavian (Saami) which has known East Asian connection..
Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
posted
Also interesting from that MA-1 paper. The researchers, insist throughout the paper, that regardless of pop culture belief, AMH occupied Eurasia DURING the LGM.
Eg Quote
15). We observe consistent results when restricting analyses to sequences with evidence of post-mortem degradation (Supplementary Information, section 15 and Supplementary Fig. 29), implying that southern Siberia may have experienced genetic continuity THROUGH the environmentally harsh LGM.
===
More proof that this Refugia retreat is also BS made up by Achilli and Torroni. Rasol and DJ was hard headed on that, time is proving me correct as usual.
-------------------- Without data you are just another person with an opinion - Deming Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
posted
Oh! To the lurkers who may not know what I am on about. In that paper by Perriria et al ? comparing mtDNA hg-U between Berbers and Saami(Scandinavia) she made a observation that the Fulani hg-U is ONE mutational step from the Saami!!!! It would interesting to see a side by side comparison between Saami and Fulani and MA-1. hg-U is one of the most ancient mtDNA lineage. Myabe close to 40,000y!!
-------------------- Without data you are just another person with an opinion - Deming Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
posted
^ One of the few instances I agree with you xy. Mind you 40,000 ya is the estimated time in which Europe was first colonized by modern humans. And yes mtDNA hg U IS associated with the paleolithic hunter-gatherers of Europe. Though the U types tha t occur in Africa tend to be of different subclades from the Eurasian ones.
-------------------- Mahirap gisingin ang nagtutulog-tulugan. Posts: 26238 | From: Atlanta, Georgia, USA | Registered: Feb 2005
| IP: Logged |
quote:Originally posted by xyyman: Oh! To the lurkers who may not know what I am on about. In that paper by Perriria et al ? comparing mtDNA hg-U between Berbers and Saami(Scandinavia) she made a observation that the Fulani hg-U is ONE mutational step from the Saami!!!! It would interesting to see a side by side comparison between Saami and Fulani and MA-1. hg-U is one of the most ancient mtDNA lineage. Myabe close to 40,000y!!
Population expansion in the North African Late Pleistocene signalled by mitochondrial DNA haplogroup U6
Intriguingly, although U5b1, which also expanded from southwest Europe in the Lateglacial, has not been seen in Moroccan Berbers, it has been identified amongst Algerian Berbers and Fulbe from Senegal, as well as Iberia, Italy and northern Eurasian Saami and Yakut [44].
Achilli A, Rengo C, Battaglia V, Pala M, Olivieri A, Fornarino S, Magri C, Scozzari R, Babudri N, Santachiara-Benerecetti AS, Bandelt H-J, Semino O, Torroni A:
Saami and Berbers - an unexpected mitochondrial DNA link. Am J Hum Genet 2005, 76:883-886
The sequencing of entire human mitochondrial DNAs belonging to haplogroup U reveals that this clade arose shortly after the out of Africa exit and rapidly radiated into numerous regionally distinct subclades. Intriguingly, the Saami of Scandinavia and the Berbers of North Africa were found to share an extremely young branch, aged merely ~9,000 years. This unexpected finding not only confirms that the Franco-Cantabrian refuge area of southwestern Europe was the source of late-glacial expansions of hunter-gatherers that repopulated northern Europe after the Last Glacial Maximum but also reveals a direct maternal link between those European hunter-gatherer populations and the Berbers.
As for U5, its sequence divergence was 8.1 ± 1.8 substitutions, corresponding to 41.4 ± 9.2 ky, a time estimate in full agreement with its proposed proto-European origin (Richards et al. 2000). It is striking that the sequence divergence of U5b1b, the subclade encompassing mtDNAs from the Saami, Yakut, Berbers, and Fulbe, was 1.7 ± 0.5 substitutions, thus corresponding to only 8.6 ± 2.4 ky.
Such a recent common ancestry of maternal lineages found in populations living as far as 9,000 miles apart and whose anthropological affinities are not at all obvious is, to say the least, unexpected. Can we provide a reasonable explanation? The recent molecular dissection of other mtDNA haplogroups reveals some clues. H1 and H3, two frequent subhaplogroups of H, display frequency peaks centered in Iberia and surrounding populations, including the Berbers of Morocco, and coalescence ages of ~11 ky (Achilli et al. 2004). Furthermore, their frequency patterns and ages resemble those reported for haplogroup V (Torroni et al. 2001a)which, similar to U5b1b, is extremely common only in the Saami (together, U5b1b and V encompass almost 90% of the Saami mtDNAs) (Torroni et al. 1996; Tambets et al. 2004). Thus, although these previous studies have highlighted the role of the Franco-Cantabrian refuge area as a major source of the hunter-gatherer populations that gradually repopulated much of central and northern Europe when climatic conditions began to improve ~15 ky ago, the identification of U5b1b now unequivocally links the maternal gene pool of the ancestral Berbers to the same refuge area and indicates that European hunter-gatherers also moved toward the south and, by crossing the Strait of Gibraltar, contributed their U5b1b, H1, H3, and V mtDNAs to modern North Africans.
Population expansion in the North African Late Pleistocene signalled by mitochondrial DNA haplogroup U6 Luísa Pereira1 2010
There is an intriguing further signal in the U6 data, witnessed by the Bayesian skyline plot. For the European haplogroup U5, which is one of the most ancient in Europe [11], we identified a strong expansion (an ~11-fold increase in effective population size) occurring in the Lateglacial period between the LGM and the beginning of the Holocene, followed by another large population expansion (~5-fold) after 5 ka, evidently associated with late Neolithic/early Bronze Age (rather than, for example, the early Neolithic expansion in Europe, which began ~8.5 ka). For U6, by contrast, the corresponding increases in effective sizes were less marked (~3-fold and ~1.5-fold, respectively), and the signal indicates that the expansion began earlier, ~22 ka. This coincides closely with the beginning of the Iberomaurusian industry in the Maghreb. These results therefore suggest that the Iberomaurusian was initiated by an expansion of modern humans of ultimately Near Eastern, carrying mtDNA haplogroup U6, who had spread into Cyrenaïca ~35-45 ka and produced the Dabban industry. The link back to the Near East and the European Early Upper Palaeolithic (which likely has the same source) may explain the suggested skeletal similarities between the robust Iberomaurusian "Mechta-Afalou" burials and European Cro-Magnon remains, as well as the case for continuity of the bearers of the Iberomaurusian industry from Morocco with later northwest African populations suggested by the dental evidence
Aside from U6, North Africa was also the recipient of European, Near Eastern and sub-Saharan African lineages most of which most likely arrived in the Holocene. Haplogroups H1, H3 and V expanded in Iberia in the Lateglacial/postglacial [11,58-61], and evidently spread into North Africa from Iberia across the Gibraltar Straits, most likely in the early Holocene [62-65]. Although the postglacial Capsian industry appears to have originated in eastern Algeria, it is tempting to hypothesize a connection with the arrival of these new populations from southwest Europe. Intriguingly, although U5b1, which also expanded from southwest Europe in the Lateglacial, has not been seen in Moroccan Berbers, it has been identified amongst Algerian Berbers and Fulbe from Senegal, as well as Iberia, Italy and northern Eurasian Saami and Yakut [44].
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
Classification of the Sami mtDNA lineages revealed that the majority are clustered in a subset of the European mtDNA pool. The two haplogroups V and U5b dominate, between them accounting for about 89% of the total. This gives the Sami regions the highest level of Haplogroups V and U5b thus far found. Both haplogroups V and U5b are spread at moderate frequencies across Europe, from Iberia to the Ural Mountains. Haplogroups H, D5 and Z represent most of the remaining averaged total. Overall 98% of the Sami mtDNA pool is encompassed within haplogroups V, U5b, H, Z, and D5. Local frequencies among the Sami vary.[3] The divergence time for the Sami haplogroup V sequences was estimated by Max Ingman and Ulf Gyllensten at 7600 YBP (years before present), and for U5b1b1 as 5500 YBP amongst Sami and 6600 YBP amongst Sami and Finns. This suggests to them an arrival in the region soon after the retreat of the glacial ice.[4] U5b
Although a small proportion of the Haplogroup U (mtDNA) among the Sami falls into U4, the great majority is U5b. The percentage of total Sami mtDNA samples tested by Tambets and her colleagues which were U5b ranged from 56.8% in Norwegian Sami to 26.5% in Swedish Sami.[3] Sami U5b falls into subclade U5b1b1. The Sami U5b1b1 [3] sub-clade is present in many different populations, e.g. 3% or higher frequencies in Karelia, Finland, and Northern-Russia.[3] The Sami U5b1 motif is additionally found in very low frequencies for instance in the Caucasus region, however this is explained as recent migration from Europe.[5] However 38% of the Sami U5b1b1 mtDNAs have haplotype so far exclusive to the Sami, containing a transition at np 16148.[3] Alessandro Achilli and colleagues noted that the Sami and the Berbers share U5b1b, which they estimated at 9,000 years old, and argued that this provides evidence for a radiation of the haplogroup from the Franco-Cantabrian refuge area of southwestern Europe.[6]
V As mentioned above, the divergence time for the Sami haplogroup V sequences was estimated by Max Ingman and Ulf Gyllensten at 7600 years ago.[4] Haplogroup V is by far the most frequent haplogroup in the Swedish Sami and is present at significantly lower frequencies in Norwegian and Finnish subpopulations.[3] Torroni and colleagues have suggested that the spread of haplogroup V in Scandinavia and in eastern Europe is due to its late Pleistocene/early Holocene expansion from a Franco-Cantabrian glacial refugium.[7] However subsequent studies found that haplogroup V is also significantly present in eastern Europeans. Furthermore, haplogroup V lineages with HVS-I transitions 16153 and 16298 that are frequent in the Sami population are much more widespread in eastern than in western Europe. So haplogroup V might have reached Fennoscandia via central/eastern Europe. Such a scenario is indirectly supported by the absence, among the Sami, of the pre-V mtDNAs that are characteristic of southwestern Europeans and northwestern Africans.[3]
Z Haplogroup Z is found at low frequency in the Sami and Northern Asian populations but is virtually absent in Europe. Several conserved substitutions group the Sami Z lineages with those from Finland and the Volga-Ural region of Russia. The estimated dating of the lineage at 2700 years suggests a small, relatively recent contribution of people from the Volga-Ural region to the Sami population.[4]
Y-DNA Three Y chromosome haplogroups dominate the distribution among the Sami: N1c (formerly N3a), I1 and R1a. The most common haplogroup among the Sami is N1c, with I1 as a close second. Haplogroup R1a in Sami is mostly seen in the Swedish Sami and Kola Sami populations, with a low level among the Finnish Sami. Tambets and colleagues suggested that N1c and R1a probably reached Fennoscandia from eastern Europe, where these haplogroups can be found in high frequencies.[3] However the two haplogroups have a distinctly different linguistic distribution. R1a1a is common among Eastern Europeans speaking Indo-European languages, while N1c correlates closely with the distribution of the Finno Ugrian languages. For example N1c is common among the Finns, while haplogroup R1a is common among all the neighbours of the Sami.[8] Haplogroup I1 is the most common haplogroup in Sweden, and the Jokkmokk Sami in Sweden have similar structure as among Swedes and Finns for haplogroup I1 and N1c.[9]
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
posted
Standing at the Gateway to Europe - The Genetic Structure of Western Balkan Populations Based on Autosomal and Haploid Markers
Kovacevic L et al.
Introduction
The Balkan Peninsula has been continuously settled by anatomically modern humans (AMH) since the Upper Paleolithic era [1][4]. The rich archaeological heritage of the region from the period of transition between Middle and Upper Paleolithics in Europe and the traces of different technologies from traditionally Neanderthal associated Mousterian to Ceramic industries of Neolithics [5][10] shows the importance of the area for understanding the spread of AMH across the continent [6],[11]. This region has been a probable gateway to Europe for first settlers [12], [13], as well as one of the refugial areas during the Last Glacial Maximum (LGM) [14], [15]. The process of the peopling of the Western Balkans a crossroad for people moving in different times to and from Europe and beyond - was extensively shaped by several historical episodes. The transition of hunting-gathering to farming in terms of the contrasting influence of pioneering agriculturalists from Anatolia and Mesolithic foragers in this area was probably complex [16], [17]. At the beginning of the second millennium BC the Balkan region was inhabited by different Illyrian tribes, which established the oldest central-western Balkan civilization [18]. The area was also the birth place of two of the world's greatest civilizations - the ancient Greek and the Byzantine Empire. The split of the Roman Empire in 395 AD divided the region into two parts, with the borderline running from Sirmium in the north (Sremska Mitrovica, Serbia) to Skadar Lake in the south (North Albania) [19]. At the same time, the Balkan region served as a frontier between the civilization of the Empire and the barbarian tribes beyond the Danube, which settled in the Balkan in the late 6th century [20], [21]. The first barbarian conquerors in the Balkans were West Goths in 410 AD [22]. In the 6th century, the Slavs had occupied the northern parts of the Danube basin and continued their way to the south. It is believed that part of the Illyrians was assimilated and the other part was forced to move south - into the territory of present-day Albania [19]. During the Great Migrations, next to the Goths and Slavs, the Mongolian tribes moved from the Central Asiatic Plateau to the Balkan Peninsula. The first of these groups of Eastern nomads to make an appearance in the Balkan were Turkic tribes: the Huns and Eurasian Avars [22], [23]. From the 15th until 19th century the Peninsula was under the Ottoman control [19], [22], [24].
Today, the Western Balkan territory (Figure 1) is inhabited by several ethnic groups of multi-religious and linguistic backgrounds. Ethnicity typically emphasizes linguistic, cultural, religious, as well as political aspects, which are human group specific, and are sometimes interpreted in different ways [25]. In this context, the term refers to religious and linguistic identity. All these groups were encompassed by the countries of the former Yugoslavian Federation and share a common recent history until 1991/1992 when a political conflict resulted in the disintegration of the Federation.
thumbnail Figure 1. Map of the Western Balkan region with triangles corresponding to the regions from where blood samples were collected.
The sample of Bosnia and Herzegovina consisted of subsamples of three main ethnic groups: Bosniacs (Sarajevo and Zavidovici), Bosnian Croats (Central Bosnia - Zepce and Maglaj; South Bosnia and Herzegovina - Mostar, Grude, Livno, Capljina), Bosnian Serbs (Doboj and Banjaluka region); Croatia (mainland, Zagreb region), Serbia (Belgrade region), Montenegro (Podgorica), Kosovo (Pristina and Prizren) and Macedonia (Skopje).
doi:10.1371/journal.pone.0105090.g001 During the last two decades the variation of uniparentally inherited markers such as mitochondrial DNA (mtDNA) and the non-recombining part of Y chromosome (NRY) have been exploited in population genetic studies in order to disentangle the problems of the diversity and dispersal of humans both in global and local contexts [26][28]. Recently, Western Balkan populations have been studied intensively from the uniparental perspective [17], [29][34]. Genetic analysis based upon the variation of Y chromosome haplogroups (hgs) has revealed that the populations of Western Balkan countries share a large fraction of the ancient gene pool of Southeastern Europe, where 70% of the paternal lineages consist of five European-specific hgs: E3b1, I-P37(xM26), J2, R1a, and R1b [31]. Marjanovic et al. [32] suggested that the frequency of NRY hg I-P37 observed in Bosnia and Herzegovina is particularly high and could be partially attributed to genetic drift. High frequencies of hg I-P37 are observed both in Bosniacs (Bosnian Muslims) (43.5%) and Bosnian Serbs (30.9%). This shows that different ethnic groups in Bosnia and Herzegovina share a large subset of their paternal lineages, affected by a major demographic event, the post-LGM expansion. A population with a high frequency of I-P37 from one of the refuges, located possibly in the Balkans, played a great role in the peopling of Bosnia and Herzegovina and surrounding areas. Similar results were observed for Croatian populations [35].
The study of the variation of mtDNA in the population of Bosnia and Herzegovina has shown - like in case of the variation of NRY - that the majority of detected mtDNA hgs among Bosnians belong to the common West Eurasian gene pool [29]. Also, it revealed that the minor part (2%) of Bosnian mtDNA lineages originate from East Eurasia and Africa. The same study observed that the differences between the Slovenian and Bosnian mtDNA pool, were likely due to two different migration waves to the Balkan Peninsula by different groups of Slavs in Middle Age [36], [37]. However, the sampled Bosnian individuals analyzed in that study were of Serbian and Croatian origin. Cvjetan et al. [30] reported that the frequencies of mtDNA hgs in populations from some countries of the former Yugoslavian Federation - Croatia (coast and mainland), Bosnia and Herzegovina, Serbia and Macedonia, including Macedonian Romani - were in concordance with Western Eurasian data. Only for the populations of small Adriatic island isolates, unusual frequencies of some mtDNA lineages have been reported which are otherwise rare in Europe [38][40]. Study of Bosch et al. [33], which included Macedonians of the former Yugoslav Republic of Macedonia, Greeks, Romanians and Albanians, as well as five Aromun populations from different parts of the Balkans, suggested that the diversity of both mtDNA and NRY hgs was similar across the Balkans, except for some Aromun populations. According to these studies, the populations of the Balkan Peninsula have been shown to be genetically homogenous and their uniparentally inherited variation is in concordance with the European genetic continuum. However, it was noted that for the better understanding of the genetic history, different intensity of mobility and migration directions of various populations of southeastern Europe, the variation of maternal lineages in the population cluster consisting of Macedonians of the former Yugoslav Republic of Macedonia, Serbians, Croatians, Herzegovinians and Bosnians should be further resolved by higher mtDNA resolution and deeper statistical analysis of sub-groups [30].
The aim of this study was to characterize, in a larger geographical context, the autosomal gene pool of eight Western Balkan populations from six countries - Bosnia and Herzegovina, Croatia, Serbia, former Yugoslav Republic of Macedonia, Montenegro and Kosovo. All studied samples were characterized also for mtDNA and NRY diversity. One of the main questions we address here is whether the whole genome approach with the accent on the variation of autosomal SNPs is in concordance with the information about genetic affinities of the populations of Western Balkan region, revealed by the studies of uniparental markers.
posted
anecdote: just 2 hours ago I was speaking (Russian) with a woman from Serbia, she quite resembled the woman's top photo except slightly wider cheeks.
What does that have to do with the thread topic about human remains in Siberia which is over 3000 miles away from Serbia from 24,000 years ago?
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
"Siberian genome reveals dual ancestry of Native Americans"
Easter Island's ancient inhabitants weren't so lonely after all
(Reuters) - They lived on a remote dot of land in the middle of the Pacific, 2,300 miles (3,700 km) west of South America and 1,100 miles (1,770 km) from the closest island, erecting huge stone figures that still stare enigmatically from the hillsides.
But the ancient Polynesian people who populated Easter Island, or Rapa Nui, were not as isolated as long believed. Scientists who conducted a genetic study, published on Thursday in the journal Current Biology, found these ancient people had significant contact with Native American populations hundreds of years before the first Westerners reached the island in 1722.
The Rapa Nui people created a unique culture best known for the 900 monumental head-and-torso stone statues known as moai erected around Easter Island. The culture flourished starting around 1200 until falling into decline by the 16th century.
Genetic data on 27 Easter Island natives indicated that interbreeding between the Rapa Nui and native people in South America occurred roughly between 1300 and 1500.
"We found evidence of gene flow between this population and Native American populations, suggesting an ancient ocean migration route between Polynesia and the Americas," said geneticist Anna-Sapfo Malaspinas of the Center for GeoGenetics at the University of Copenhagen, who led the study.
The genetic evidence indicates either that Rapa Nui people traveled to South America or that Native Americans journeyed to Easter Island. The researchers said it probably was the Rapa Nui people making the arduous ocean round trips.
"It seems most likely that they voyaged from Rapa Nui to South America and brought South Americans back to Rapa Nui and admixed with them," said Mark Stoneking, a geneticist with Germany's Max Planck Institute for Evolutionary Anthropology who collaborated on a related study of Brazil's indigenous Botocudo people. "So it will be interesting to see if in further studies any signal of Polynesian, Rapa Nui ancestry can be found in South Americans."
In making their way to South America and back, the Rapa Nui people may have spent perilous weeks in wooden outrigger canoes.
The researchers concluded that the intermixing occurred 19 to 23 generations ago. They said Rapa Nui people are not believed to have started mixing with Europeans until much later, the 19th century. Malaspinas said the genetic ancestry of today's Rapa Nui people is roughly 75 percent Polynesian, 15 percent European and 10 percent Native American.
A second study, also published in Thursday's issue of Current Biology, illustrates another case of Polynesians venturing into South America. Two ancient human skulls from Brazil's indigenous Botocudo people, known for the large wooden disks they wore in their lips and ears, belonged to people who were genetically Polynesian, with no detectable Native American ancestry.
"How the two Polynesian individuals belonging to the Botocudos came into Brazil is the million-dollar question," said University of Copenhagen geneticist Eske Willerslev of the Center for GeoGenetics, who led the study on the Botocudos.
The findings suggest these Polynesians reached South America and made their way to Brazil, either landing on the western coast of the continent and crossing the interior or voyaging around Tierra del Fuego and up the east coast, Stoneking said.
"In either event it is an amazing story," he said.
Nature 514, 445449 (23 October 2014) doi:10.1038/nature13810
Genome sequence of a 45,000-year-old modern human from western Siberia
Qiaomei Fu et al.
We present the high-quality genome sequence of a ~45,000-year-old modern human male from Siberia. This individual derives from a population that lived beforeor simultaneously withthe separation of the populations in western and eastern Eurasia and carries a similar amount of Neanderthal ancestry as present-day Eurasians. However, the genomic segments of Neanderthal ancestry are substantially longer than those observed in present-day individuals, indicating that Neanderthal gene flow into the ancestors of this individual occurred 7,00013,000 years before he lived. We estimate an autosomal mutation rate of 0.4 × 10−9 to 0.6 × 10−9 per site per year, a Y chromosomal mutation rate of 0.7 × 10−9 to 0.9 × 10−9 per site per year based on the additional substitutions that have occurred in present-day non-Africans compared to this genome, and a mitochondrial mutation rate of 1.8 × 10−8 to 3.2 × 10−8 per site per year based on the age of the bone.
Y DNA K(xLT
(Supp, section 9)
mtDNA Ust-Ishim
The Ust-Ishim mtDNA sequence falls at the root of a large group of related mtDNAs (the R haplogroup), which occurs today across Eurasia (Supplementary Information section 8). It is clear that this was a Eurasian individual: Based on genotyping data for 87 African and 108 non-African individuals (Supplementary Information section 11), the Ust-Ishim genome shares more alleles with non-Africans than with sub-Saharan Africans (|Z| = 4189), consistent with the principal component analysis, mtDNA and Y chromosome results.
Among the non-Africans, the Ust-Ishim genome shares more derived alleles with present-day people from East Asia than with present-day Europeans (|Z| = 2.16.4).
However, when an ~8,000-year-old genome from western Europe (La Braña)9 or a 24,000-year-old genome from Siberia (Malta 1)10 were analysed, there is no evidence that the Ust-Ishim genome shares more derived alleles with present-day East Asians than with these prehistoric individuals (|Z| < 2). This suggests that the population to which the Ust-Ishim individual belonged diverged from the ancestors of present-day West Eurasian and East Eurasian populations beforeor simultaneously withtheir divergence from each other. The finding that the Ust-Ishim individual is equally closely related to present-day Asians and to 8,000- to 24,000-year-old individuals from western Eurasia, but not to present-day Europeans, is compatible with the hypothesis that present-day Europeans derive some of their ancestry from a population that did not participate in the initial dispersals of modern humans into Europe and Asia11.
we estimate that the admixture between the ancestors of the Ust-Ishim individual and Neanderthals occurred approximately 50,000 to 60,000 years BP, which is close to the time of the major expansion of modern humans out of Africa and the Middle East.
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
posted
Did you all think I was joking when I said all Xyyman posts to EGYPTOLOGY (and any replies to them) will be deleted for the unforeseeable future due to his continued use of hateful epithets?
You all can't have it both ways.
If you all really want a moderator (as something other than a flunky to handle your personal booboos) to raise this forum a level or two then why don't you abide by what the moderator does to improve ES?
Posts: 8675 | From: Tukuler al~Takruri as Ardo since OCT2014 | Registered: Feb 2003
| IP: Logged |






 UBBFriend: Email this page to someone!
UBBFriend: Email this page to someone!











![[Roll Eyes]](rolleyes.gif)





 Printer-friendly view of this topic
Printer-friendly view of this topic