Introduction Probably the most striking finding derived from the Neandertal genome project [1] was the evidence for admixture between Neandertals and a population of modern humans that left Africa between 80 Kya and 50 Kya subsequently expanding into the rest of the world. The study involved the sequencing and comparison of the Neandertal genome to five modern human genomes: two African (Yoruba and San) and three non-Africans (French, Chinese and Melanesian); all the non-African human genomes shared with Neandertals between 14% of their genome, in regions of low recombination placed along ten chromosomes [1]. Additional genomic region introgressions from Neandertals, Denisovans and also putative archaic African hominins have been recently described in Eurasian, Oceanic and even African populations, respectively [2][7].
However, an alternative scenario in which the ancestral population of today non-Africans was more closely related to Neandertals than the ancestral population of current Africans due to ancient substructure within the African continent, cannot be totally excluded with the present data [8], although it seems unlikely [9]. In light of this, it is unfortunate that North African individuals have not been included in these admixture analyses, since both the putative African substructure and the admixture are likely to differentially affect North African and sub-Saharan African populations.
The importance of North Africa in the emergence of modern Homo sapiens has been traditionally neglected. However, recent archaeological and paleontological evidence increasingly points to this area as a potential source of out-of-Africa migrations [10],[11]. Recent dating of the characteristic North African lithic industry, called Aterian, has provided much older dates than previously assumed, now ranging from 145 Kya to 40 Kya [12],[13]. These Aterian people made personal ornaments with shells, a sign of modern human symbolic behavior [14]. Morphometric analyses of the 80 Kya Dar es-Soltan skull (Morocco) and of Aterian hominin teeth show similarities with early modern humans from Qafzeh and Skhul (Israel) and with the later skull of Pestera cu Oase (Romania) [15],[16].
Recent genetic analysis of North African populations [17] have found that, despite the complex admixture genetic background, there is an autochthonous genomic component which is likely derived from back-to-Africa gene flow older than 12,000 years ago (ya) (i.e., prior to the Neolithic migrations). This local population substratum seems to represent a genetic discontinuity with the earliest modern human settlers of North Africa (those with the Aterian industry) given the estimated ancestry is younger than 40,000 years ago [17]. The estimated time of Neandertal admixture with modern human populations is between 37,00086,000 years ago [18].
The aim of this work was to investigate if this autochthonous North African ancestry bares any traces of the introgression with Neandertals, by applying the f4 ancestry ratio statistic test, previously used to detect Denisovan admixture in Asia [3]. We show that North African populations, like all non-African humans [1], also carry the signature of admixture with Neandertals, and that the real geographical limit for Neandertal admixture is between sub-Saharan groups and the rest.
Materials and Methods Top To ascertain whether or not current North African populations show any signs of Neandertal admixture, we analyzed recently published data of 125 North African individuals genotyped with the Affymetrix 6.0 chip and accounting for 780,000 SNPs were analyzed [17]. Individuals are representative of seven different North African locations (Table 1) spanning from west to east. To have a broader coverage of Eurasia and to allow comparison with Sub-Saharan populations, African and Eurasian populations were included in the analysis [17],[19],[20].
In order to compare the human SNP data to the Neandertal, bam read files from all Neandertal samples from the UCSC ftp site (ftp://hgdownload.cse.ucsc.edu/gbdb/hg18/neandertal/seqAlis) were downloaded and merged. Base and mapping quality filters reported in previous studies were implemented in the analysis [2],[21]. To avoid any confusion with ancient DNA postmortem modifications, C-T and G-A human ancient hominin nucleotide sites, were discarded. For all sequencing data, a single read was randomly sampled for each individual at positions overlapping the array SNPs coordinates. Furthermore all human and Neandertal data were merged with sequence data from chimpanzee (CGSC 2.1/Pantro), and data were further processed to control for strand misidentification [3], to conform a final data set of 142,720 SNPs.
North African populations have a complex genetic background. In addition to an autochthonous genetic component, they exhibit signals of European, sub-Saharan and Near Eastern admixture as previously described [17]. Moreover, the use of genotype data can suffer from potential biases that arise from discovering SNPs in a limited number of individuals, thus resulting in enrichment of common alleles, particularly in the populations from which the discovery panel was constructed [22],[23] (in the present case would be a bias towards European populations). Two challenges arise from these effects: first, patterns of gene flow detected between Neandertal and North Africans could be the consequence of subsequent admixture between North Africans and other modern human populations and second, the ascertainment bias towards European and East Asian populations could magnify differences in signals of Neandertal gene flow in individuals with high Sub-Saharan ancestry compared to individuals with high European ancestry.
In order to overcome these problems we initially assessed the different genetic components in North African populations using an unsupervised clustering algorithm, ADMIXTURE [24], on a sample set of around 50,000 SNPs that included all North African individuals, together with populations of European, Near Eastern and Sub-Saharan origin [17],[25],[26].
As a first approach to establish the relationship between North African populations and Neandertal, a projected Principal Component Analysis (PCA) was performed. In addition to the chimpanzee and the Neandertal genomes, data from the Denisova genome were downloaded and merged in this case resulting in 111,991 SNPs (after filtering for strand bias SNPs and ancient DNA miscoding lesions). Given that the ancient hominin and chimpanzee genomes have been originally sequenced at low coverage no SNP polymorphism data are available, and therefore individuals were considered at the haplotype level only. First, a PCA was generated using Neandertal, chimpanzee, and Denisova. Then, SNP loadings for the first two components were used to project the sample set of modern humans.
Next, we aimed at estimating the amount of Neandertal admixture in North African populations using the f4 ancestry ratio test [27]. Although a previous simulation study [28] suggested that the analysis of SNP data from arrays can provide biased results in admixture estimates, there is more recent evidence supporting that f4 ancestry ratio statistic is unaffected by those biases [3]. The f4 ancestry ratio test measures the proportion of archaic hominin genetic fraction in a modern human population as a fraction of the known amount of archaic introgression in another modern human population. Consequently, the f4 ancestry ratio test basically measures the correlation in allele frequency differences between two populations used as outgroups (e.g., chimpanzee and Neandertal), a Sub-Saharan African population (Yorubans) and the X-tested population, normalized by the correlation in allele frequency differences between chimpanzee, Neandertal, a Sub-Saharan African group (Yorubans) and a human population previously known to have experienced Neandertal admixture (in this case, CEU) [1]. If Yorubans and X descend from a single ancestral population without any subsequent admixture with Neandertals, then the allele frequency differences between Yorubans and X must have arisen solely after their separation from their common ancestor; the two frequency differences should be uncorrelated and thus the f4 ancestry ratio statistic should have an expected value of zero.
Finally, a block jackknife [29],[30] approach was used to estimate standard errors; blocks were separated by dropping each non-overlapping five cM stretch of the genome in turn, and studying the variance of each statistic of interest to obtain a approximately normal distributed standard error [25]. Further combinations (e.g. San instead of Yoruban and Chinese instead of CEU) were also calculated to test the consistency of the results (Table S1).
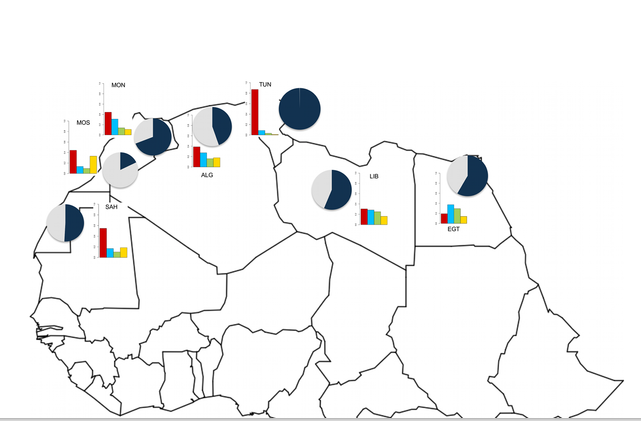
Results and Discussion Top We ran ADMIXTURE for k equal 2 to 7 and obtained CV errors, and determined that the best k (the one with lowest cross-validation error) is k = 4. Results (Figure 1) are coincident with those previously published [17] and show that North Morocco, Libya and Egypt carry high proportions of European and Near Eastern ancestral components, whereas Tunisian Berbers and Saharawi are those populations with highest autochthonous North African component. Particularly, ten Tunisian individuals have more than 99% of their genome assigned to North African ancestry and therefore have been analyzed separately (subsequently referred to as N-TUN) from the overall Tunisian population.
In the PCA analysis (Figure 2) Eurasian populations are the closest to Neandertals among modern humans, which is in agreement with previous studies [1]. Sub-Saharan Africans are, as expected, more distant to Neandertal, whereas North African individuals are placed between these two groups. North African individuals with the highest Sub-Saharan African component (as detected by ADMIXTURE) are distant from Neandertal and closer to Sub-Saharan populations. It is interesting to notice that the North African populations closer to Neandertals are populations with a large known European or Near Eastern admixture, but also the Tunisians that have an almost complete autochthonous North African genetic component.
The results of the f4 ancestry ratio test (Table 2 and Table S1) show that North African populations vary in the percentage of Neandertal inferred admixture, primarily depending on the amount of European or Near Eastern ancestry they present (Table 1). Populations like North Morocco and Egypt, with the highest European and Near Eastern component (~40%), have also the highest amount of Neandertal ancestry (~6070%) (Figure 3). On the contrary, South Morocco that exhibits the highest Sub-Saharan component (~60%), shows the lowest Neandertal signal (20%). Interestingly, the analysis of the Tunisian and N-TUN populations shows a higher Neandertal ancestry component than any other North African population and at least the same (or even higher) as other Eurasian populations (100138%) (Figure 3).
Some results of the estimated ancestry in Table 2 are higher than 100%. Because the amount of Neandertal admixture provided by this statistic is in relation to the fraction found in another population, populations with more than 100% values, have more than the observed Neandertal admixture levels found in the source population used for comparison (i.e CEU). On the other hand, a negative f4 ancestry ratio value such as that one observed for the Luyha in Table 2 could have several explanations. One possibility is that it reflects an artifact of ascertainment bias on SNP arrays. Ascertainment bias is likely to affect the joint information from Europeans and East Asians, since SNP arrays are most commonly designed based on information from these populations. On the other hand it could also reflect a more complex demographic history (i.e population structure between the populations being compared) than previously assumed.
Subsequently, we aimed to compare the results revealed by ADMIXTURE and by the f4 ancestry ratio statistic in an attempt to corroborate that the signal of Neandertal admixture revealed in North African populations is not caused by Eurasian admixture. For this purpose, we performed a Pearson correlation test between the ancestry proportions estimated with ADMIXTURE and the proportions of Neandertal admixture estimated by the f4 ancestry ratio test. Specifically, we tested the correlation between a) both European and Near Eastern components and Neandertal admixture and b) European, Near Eastern and North African admixture components and Neandertal admixture. If signals of gene flow from Neandertals were due exclusively to the European and the Near Eastern components, we would expect that the correlation should significantly decrease in test b), when the North African component is included. On the contrary, the Pearson correlation test reaches significance only when the North African component is included, which is maintained even when Tunisians are removed from the analysis (Table 3).
Overall, the correlation analysis and the f4 ancestry ratio statistic show that the North African component actually contributes to the signal of gene flow from Neandertals. Given that the North African autochthonous ancestry seems to be 12,00040,000 years old [17], our hypothesis is that this ancestral population was descendant from the populations that first interbreed with Neandertals about ~37,00086,000 years ago [18] somewhere in the Middle East. Nonetheless further analyses in populations around the contact areas are needed to confirm this hypothesis.
A previous study [26] observed that the similarity to Neandertals increases with distance from Africa and suggested this could be explained by SNP ascertainment bias plus a strong genetic drift in East Asian populations. Nonetheless more complex, population-biased, ascertainment schemes might have additional effects (i.e bottlenecks), but these are not expected to significantly increase the rate of false positives in admixture tests [31]. The Tunisian population has been reported to be a genetic isolate [17] so it is plausible that part of the signal detected is actually due to genetic drift. However, this should not affect the other North African groups in our study. Finally, given that SNP arrays are based on common alleles and probably the relevant admixture information is encoded within the rare and very rare alleles, the potential bias, if anything, will underestimate ancient hominid admixture signals, as shown in previous studies [2],[3].
With the current data, however, it is not possible to discard the ancient African substructure hypothesis [8]. Although ours and some previous results [9] tend to favor the admixture hypothesis as the most plausible one, we think that a complete clarification of this issue can only be achieved with a Neandertal high coverage genome, such as this recently achieved for Denisova [32]. This, and sequencing data of North African populations, especially those with a high autochthonous component, may help elucidate more precisely the interbreeding process with Neandertals. In any case, our results show that Neandertal genomic traces do not mark a division between African and non-Africans but rather a division between Sub-Saharan Africans and the rest of modern human groups, including those from North Africa.
Supporting Information Top Table S1.
Stability of the Neandertal admixture estimates. We present each population's estimate ancestry, the standard error in the estimate, and the Z score for different combinations of Sub-Saharan and non-African populations. O (Out-group), BP (Benchmark population, i.e. population which didn't experience any introgression from Neandertals) and SP (Source population i.e. populations in which the amount of introgression from Neandertal is known).
Posts: 42940 | From: , | Registered: Jan 2010
| IP: Logged |
posted
^^Some other scholars express skepticism towards claims of Neanderthal admixture as shown below
Abstract "Recent comparisons between anatomically modern humans and ancient genomes of other hominins have raised the tantalizing, and hotly debated, possibility of hybridization. Although several tests of hybridization have been devised, they all rely on the degree to which different modern populations share genetic polymorphisms with the ancient genomes of other hominins. However, spatial population structure is expected to generate genetic patterns similar to those that might be attributed to hybridization. To investigate this problem, we take Neanderthals as a case study, and build a spatially explicit model of the shared history of anatomically modern humans and this hominin. We show that the excess polymorphism shared between Eurasians and Neanderthals is compatible with scenarios in which no hybridization occurred, and is strongly linked to the strength of population structure in ancient populations. Thus, we recommend caution in inferring admixture from geographic patterns of shared polymorphisms, and argue that future attempts to investigate ancient hybridization between humans and other hominins should explicitly account for population structure." --Anders Eriksson, Andrea Manica (2012) Effect of ancient population structure on the degree of polymorphism shared between modern human populations and ancient hominins. PNAS 2012
---------------------------------------------- New research raises doubts about whether modern humans and Neanderthals interbred August 13, 2012
New research raises questions about the theory that modern humans and Neanderthals at some point interbred, known as hybridisation. The findings of a study by researchers at the University of Cambridge suggests that common ancestry, not hybridisation, better explains the average 1-4 per cent DNA that those of European and Asian descent (Eurasians) share with Neanderthals. It was published today, 13 August, in the journal PNAS.
In the last two years, a number of studies have suggested that modern humans and Neanderthals had at some point interbred. Genetic evidence shows that on average Eurasians and Neanderthals share between 1-4 per cent of their DNA.
In contrast, Africans have almost none of the Neanderthal genome. The previous studies concluded that these differences could be explained by hybridisation which occurred as modern humans exited Africa and bred with the Neanderthals who already inhabited Europe. However, a new study funded by the BBSRC and the Leverhulme Trust has provided an alternative explanation for the genetic similarities. The scientists found that common ancestry, without any hybridis"ation, explains the genetic similarities between Neanderthals and modern humans.
In other words, the DNA that Neanderthal and modern humans share can all be attributed to their common origin, without any recent influx of Neanderthal DNA into modern humans. Dr Andrea Manica, from the University of Cambridge, who led the study said: "Our work shows clearly that the patterns currently seen in the Neanderthal genome are not exceptional, and are in line with our expectations of what we would see without hybridisation. So, if any hybridisation happened - it's difficult to conclusively prove it never happened - then it would have been minimal and much less than what people are claiming now."
Neanderthals and modern humans once shared a common ancestor who is thought to have spanned Africa and Europe about half a million years ago. Just as there are very different populations across Europe today, populations of that common ancestor would not have been completely mixed across continents, but rather closer populations would have been more genetically similar to each other than populations further apart. (There is extensive genetic and archaeological evidence that population in Africa were 'structured'; in other words, different populations in Africa only had limited exchange through migration, allowing them to remain distinct from each other both in terms of genetics and morphology.)
Then, about 350-300 thousand years ago, the European range and the African range became separated. The European range evolved into Neanderthal, the African range eventually turned into modern humans. However, because the populations within each continent were not freely mixing, the DNA of the modern human population in Africa that were ancestrally closer to Europe would have retained more of the ancestral DNA (specifically, genetic variants) that is also shared with Neanderthals. On this basis, the scientists created a model to determine whether the differences in genetic similarities with Neanderthal among modern human populations, which had been attributed to hybridisation, could be down to the proximity of modern humans in northern Africa (who would have later gone on to populate Europe) to Neanderthals.
By examining the different genetic makeup among modern human populations, the scientists' model was able to infer how much genetic similarity there would have been between distinct populations within a continent. The researchers then simulated a large number of populations representing Africa and Eurasia over the last half a million years, and estimated how much similarity would be expected between a random Neanderthal individual and modern humans in Africa and Eurasia.
The scientists concluded that when modern humans expanded out of Africa 60-70K years ago, they would have brought out that additional genetic similarity with them, making Europeans and Asians more similar to Neanderthals than Africans are on average undermining the theory that hybridization, and not common ancestry, explained these differences.
Dr Manica added: "Thus, based on common ancestry and geographic differences among populations within each continent, we would predict out of Africa populations to be more similar to Neanderthals than their African counterparts - exactly the patterns that were observed when the Neanderthal genome was sequenced; but this pattern was attributed to hybridisation. Hopefully, everyone will become more cautious before invoking hybridisation, and start taking into account that ancient populations differed from each other probably as much as modern populations do."
More information: Effect of ancient population structure on the degree of polymorphism shared between modern human populations and ancient hominins, PNAS, August 13, 2012. Journal reference: Proceedings of the National Academy of Sciences Provided by University of Cambridge
DATA RECAP: 2010 Berber mtDNA study finds Berber roots foundational in Africa - frigi 2010
Ancient Local Evolution of African mtDNA Haplogroups in Tunisian Berber Populations
Frigi et al.
Human Biology
August 2010 (82:4)
Discussion In this study we attempted to better elucidate the ancient African genetic background in the northwest African area, particularly in Tunisia. To this aim, we focused our study on Berber populations that are considered representative of the ancient North African populations that probably derived from Neolithic Capsians. During historic times, Berbers experienced a long and complicated history with many invasions, conquests, and migrations by Phoenicians, Romans, Vandals, Byzantines, Arabs, Bedouins, Spanish, Turks, Andalusians, sub-Saharans (communities settled in Jerba and Gabes in the 16th19th centuries), and French (Brett and Fentress 1996). During these invasions, Berbers were forced back to the mountains and to certain villages in southern Tunisia (Fadhlaoui-Zid et al. 2004). At present, they are restricted to some isolates in the south who maintain the Berber language and to some populations in the north who lack an origin language. Many genetic studies on Tunisian Berber populations demonstrate the heterogeneity of Berbers with respect to European and sub-Saharan African contributions and the mosaic structure of Tunisian Berber populations with an absence of ethnic, linguistic, and geographic effects (Cherni et al. 2010).
In the present work, mtDNA data show a diversified distribution of African haplogroups. However, a question remains concerning the date of the sub-Saharan African inputs. Our results demonstrate an ancient local evolution in Tunisia of some African haplogroups (L2a, L3*, and L3b). The most ancient haplogroup is L3*, which would have been introduced from eastern sub-Saharan populations to North Africa about 20,000 years ago. The Siwa oasis sample studied by Coudray et al. (2009) contains sub-Saharan haplogroups L0a1, L3i, L4*, and L4b2, which are different from our Tunisian samples, in agreement with the heterogeneity of Berbers already shown in Tunisia.
Stevanovitch et al. (2004) suggested that the Gurna population in Egypt has conserved the trace of an ancestral genetic structure from an ancestral East African population characterized by a high haplogroup M1 frequency. This haplogroup is also present in three Berber populations (Kesra, Matmata, and Sned) with variable frequencies. In each of these populations, haplogroup L3* is also present. The association of both eastern African haplogroups in the Berber populations is a strong argument in favor of eastern African gene flow in Berbers. Other genetic and archaeological studies confirmed the crucial idea that an ancient population in East Africa constituted the basis of the ancestors of all African Upper Paleolithic populationsand their subsequent present-day descendants (Bengtson 2008; Keita 2004; Relethford 2000; Zakrzewski 2003, 2007).
Moreover, Berber languages spoken exclusively by North African populations belong to the Afro-Asiatic language. Diakonoff (1998) showed an exclusively African origin (Diakonoff, 1981, 1988) for the family. He explicitly described proto-Afro-Asiatic vocabulary as consistent with non-food-producing vocabulary and linked it to pre-Neolithic cultures in the Levant and in Africa south of Egypt. Moreover, Ehret. (2003) suggested that early Afro-Asiatic languages were spread by Mesolithic foragers from Africa into the Levant. On the contrary, Diamond and Bellwood (2003) suggested that food production and the Afro-Asiatic language family were brought simultaneously from the Near East to Africa by demic diffusionin other words, by a migration of food-producing peoples. The evidence presented by Wetterstrom (1993) does not support this latter suggestion, however, and indicates that early African farmers in the Fayum initially incorporated Near Eastern domesticates into an existing indigenous foraging strategy and only over time developed a dependence on horticulture.
In conclusion, the crucial linguistic finding is that the three deepest clades of the Afro-Asiatic family are localized in Eritrea and Ethiopia. All the other languages of the family outside that region belong to subclades of just one of those deep clades. This kind of cladistic distribution is a basic criterion of the genetic argument for the genetic lineage origins well understood by geneticists. It applies to linguistic history as well.
Our results also point to a less ancient western African gene flow to Tunisia involving haplogroups L2a and L3b. Thus the sub-Saharan contribution to northern Africa starting from the east would have taken place before the Neolithic. The western African contribution to North Africa should have occurred before the Saharas formation (15,000 years BP). It seems likely that an expansion would have taken place in the Sahel zone starting about the time of a gradual climatic return to wetter conditions, when the Senegal River cut through the dunes (Burke et al. 1971). For subhaplogroup L2a1 (data not shown) we found some haplotypes that the Tunisian Berbers shared with Mauritanians and western sub-Saharan populations speaking a Niger-Congo language (studied by Salas et al. 2002).
This suggests that the people who brought these markers to the Berber populations most likely came from West African populations that spoke languages belonging to the Niger-Congo family when the Sahara became drier. However, this contribution of West African haplotypes and of other haplotypes, such as those belonging to haplogroup L1b1, could have been introduced to North Africa more recently.
Indeed, this West African contribution was difficult to date, because few haplotypes belonging to western African haplogroups have been observed, most of them being divergent. This result can be interpreted in different ways. Ancient western African mtDNA contributions could have disappeared from North Africa as a result of recent flows, or the situation observed now could be the result of a strong drift effect on ancient western African lineages, particularly those belonging to haplogroups L2a and L3b. A strong Iberian gene flow may have contributed to the decrease in African haplogroups. Indeed, most of the older hypotheses about North African population settlement used to suppose an Iberian or an eastern origin. The dates for subhaplogroups H1 and H3 (13,000 and 10,000 years, respectively) in Iberian and North African populations allow for this possibility. Kefi et al.s (2005) data on ancient DNA could be viewed as being in agreement with such a presence in North Africa in ancient times (about 15,0006,000 years ago) and with the fact that the North African populations are considered by most scholars as having their closest relations with European and Asian populations (Cherni et al. 2008; Ennafaa et al. 2009; Kefi et al. 2005; Rando et al. 1998).
However, considering the general understanding nowadays that human settlement of the rest of the world emerged from eastern northern Africa less than 50,000 years ago, a better explanation of these haplogroups might be that their frequencies reflect the original modern human population of these parts of Africa as much as or more than intrusions from outside the continent. The ways that gene frequencies may increase or decrease based on adaptive selection, gene flow, and/or social processes is under study and would benefit from the results of studies on autosomal and Y-chromosome markers.
Since the end of the extreme Saharan desiccation, lasting from before 25,000 years ago up to about 15,000 years ago, the Sahara has had post- and pre- Holocene cyclical climatic changes (Street and Grove 1976), and corresponding increases and decreases in population are probable. Wetter phases with better habitats perhaps allowed for increased colonization and gene and cultural exchange. Desiccation would have encouraged the emigration and segmentation of populations, with resultant genetic consequences secondary to drift producing more variation. During the last glacial period, the Sahara was even bigger than it is today, extending south beyond its current boundaries (Ehret 2002). About 13,000 years ago, large parts of the Sahara were as dry as the desert is now (White and Mattingly 2006). The end of the glacial period brought more rain to the Sahara, especially from about 8500 to 6000 BC (Fezzan Project 2006). By around 3400 BC, the monsoon retreated south to approximately where it is today, leading to the gradual desertification of the region (Kröpelin 2008). Thus the Sahara, through its cyclical environmental changes, might be seen as a microevolutionary processor and/or pump of African people that ejected groups to the circum-Saharan regions in times of increasing aridity.
Indeed, it must be noted that the high frequencies of cDe, P, and V antigens and low frequencies of FY antigens in some Berber-speaking groups (Chamla 1980; Mourant et al. 1976) indicate affinities with tropical Africans. These data may indicate recent or ancient gene flow from sub-Saharan Africa, a common immediate pre-Holocene ancestral group, or chance resemblance.
Our findings are in accordance with other studies on Y-chromosome markers that have shown that the predominant Y-chromosome lineage in Berber communities is the subhaplogroup E1b1b1b (E-M81), which emerged in Africa, is specific to North African populations, and is almost absent in Europe, except in Iberia (Spain and Portugal) and Sicily. Molecular studies on the Y chromosome in North Africa are interpreted as indicating that the southern part of Africa, namely, the Horn/East Africa, was a major source of population in the Nile Valley and northwest Africa after the Last Glacial Maximum, with some migration into the Near East and southern Europe (Bosch et al. 2001; Underhill et al. 2001). Hence, contrary to the suggestion that mtDNA haplogroups were introduced mostly from Iberia, it seems that Y-chromosome markers have an eastern African origin with an ancient local evolution in North Africa. These observations are in agreement with the proposal that the ancient communities ancestral in language to more recent Berber communities absorbed a lot of females from the existing pre-Holocene populations. This would indicate that the North African populations arose from admixture rather than from local evolution, leading to an intermediate genetic structure between eastern sub-Saharan Africans and Eurasians. Rock paintings in North Africa that show people of different phenotypes living together are a strong argument for our hypothesis (Hachid 1982, 1992, 1998).
In conclusion, our findings parallel the more recent findings of both archaeology and linguistics on the prehistory of Africa. The present study suggests that sub-Saharan contributions to North Africa have experienced several complex population processes after the occupation of the region by anatomically modern humans. Our results reveal that Berber speakers have a foundational biogeographic root in Africa and that deep African lineages have continued to evolve in supra-Saharan Africa.
Biocultural Emergence of the Amazigh in Africa: Comment on Frigi et al. (2010) S. O. Y. Keita. Human Biology (August 2010) (82:4)
Frigi et al. [2010 (this issue)] present some new findings on a population of AmazighBerber speakers in Tunisia. Although their study is not exhaustive, they provide an outline of human population history in the Maghreb and a general discussion of its mtDNA diversity. Their work is important in inviting researchers to think about the concepts of continuity and change in biology, culture, language, and identity in a geographic space. Their presentation helps in understanding the complexity of examining the ancestry and emergence of Berber origins in Africa as a local process and encourages the consideration of many questions about how the human biology and culture of a known population or ethnolinguistic group can be conceptualized through space and time.
Berber- (Tamazight-) speaking communities are thought to represent the clearest known descendants of the ancient indigenous populations of Africa west of the northern Nile valley in the supra-Saharan and northern Saharan regions (Brett and Fentress 1996; Camps 1982; Desanges 1981). Indigenous here can refer only to those whom we can perceive as having had the longest tenure on the land, using available historical evidence. However, there are questions. What constitutes historical evidence for earlier periods? Should it include archaeology, paleontology, historical linguistics, skeletal biology, and genetics, as broadly advocated by a historical anthropological approach (e.g., Kirch and Green 2001; Mace et al. 2005)? Or is it only to be based on the interpretation of texts from the ancient Egyptian, Greco-Roman, or Islamic periods [e.g., see comments by Brett and Fentress (1996), Desanges (1981), Norris 1982, and Snowden (1971)]?
How is the varied evidence to be ranked in importance, reconciled when it seems to be in contradiction, and analyzed synthetically? A simplistic positivism has to be avoided in discussions of any facet of human history because various pathways could lead to similar results. What is the role of evolutionary mechanismsadaptive selection, gene flow, drift, sexual selectionin explaining the biology of some of the ancestors of Berber speakers at the deepest time levels and of living Amazigh as well? Frigi et al. (2010) do not address all these questions directly, but their work implicitly acknowledges their importance and provides a new framework for investigation.
Braudels (1980) concept of different levels of history can be adapted and adopted in a modified form as levels of biocultural or bioethnic history, to further consider Frigi and colleagues contribution, which implicitly acknowledges the contingent and multidimensional character of population interactions through time against an evolutionary background.
Another issue of some interest is the (mis)labeling of Berbers as Eurasian migrants from the Near East: Did they arrive as a unit from Asia or Europe, as a settler colonist package with a persistent identity analogous to Europeans in South Africa, or did the biology, language, and culture of the Amazigh emerge primarily from a set of interactions in Africa involving African peoples at base, that is, as a part of authentic African historical and biological processes? There are no ancient Berber communities outside Africa, and the idea of simple demic diffusion of Berbers as a people to the Maghreb (e.g., Arredi et al. 2004) from the Near East is not supported. It is of some interest that even Coon and Hunt (1965), using a raciotypological paradigm now long discredited, postulated a massive invasion of Africa by Caucasians in the Pleistocene and therefore thought that Berber language and identity had entered the Maghreb from more southerly regions in Africa. Frigi and colleagues suggest that several populations over time were involved in the biological ancestry of the current Berber speakers, and this is consistent with archaeological evidence of actual migration in the mid- to late Holocene (Camps 1982) as well as historical documentation. Craniofacial diversity has been documented in the region before Vandal and Arab migrations (Keita 1990).
It is important to remember that biology, language, and culture are not intrinsically or obligatorily correlated, a principle established some time ago (Boas 1940). It is not particularly surprising that one can sometimes find markers that will correlate across biology and culture at some levelsbut the issue is how this came to be and when, and what it represents historically and socially. There is always the question of whether the correlation is an artifact of recent events or of primordial ontology. A language or language family can be adopted slowly or rapidly by nonnative speakers. The current biological profile of a region may predate (or postdate) the language spoken there. Communities may adopt (nonlinguistic) cultural practices from others without greatly changing language or biology; or they may become primarily integrated linguistically and politically but not biologically or exhibit other permutations of these variables, such as total biocultural assimilation. The biology of a particular ethnolinguistic group or community may change based on intermarriage if the social rules allow the offspring to become group members. Such matings may have occurred long before the recall of communal memory, whether in texts or oral tradition. Relatively nonethnocentric polygamous societies or populations may have cultural descendants who are genealogically heterogeneous when viewed over a millennium.
Frigi et al. (2010) suggest these possibilities as factors in their consideration of the asymmetric assimilation of females of non-African origin into Berber-speaking populations whose males currently have a predominance of lineages defined by the African M35/81 biallelic marker. It is interesting that these non-African mtDNA lineages are usually predominant while being diverse (Coudray et al. 2009; Fadhlaoui-Zid et al. 2004; Khodjet-el-Khil et al. 2008). The existence of mtDNA lineages common to Saami and some Amazigh groups (Achilli et al.
2005) is likely to be explained by the migration of females bearing these lineages from a region in northern Europe (perhaps in the ranks of the Vandals or far more ancient back-migrations to Africa), whether they were ethnically Saami or not. However, there may have been other locales where these lineages once existed. Circular reasoning in syntheses involving multiple disciplines has to be avoided. The criteria and methods for a given discipline usually have to be given equal weight, and their results should be considered independently before an effort at synthesis is made. For example, a hypothesis about the place of origin of a language family or phylum must be based on linguistic evidence and methods, not on DNA or craniofacial patterns. Likewise the place of origin of a particular genetic variant or lineage has to be based on genetic data, principles, and models, not on archaeological data. The locale of origin of a particular culture or archaeological industry is subject to analyses based on methods and theory that are specific to the relevant disciplines.
The only exception to these rules is if a calculated date of origin of a genetic variant found in a given locale predates the existence of people in that place. Although the notion of population ties together both biology and culture broadly conceived, it cannot be claimed that continuity in one necessarily means continuity in another. If the question is about physical population migration, then the same conclusion reached from every discipline independently would seem to best support the claim (Rouse 1986). However, it cannot be said absolutely that there was no movement if all lines of evidence do not point in the same direction. The idea of Occams razor may sometimes mean accepting the reality of human complexity and an inability to reconcile evidence with preconceived models.
Frigi et al. (2010) have provided a general temporal framework for Maghreb population history, from the Paleolithic to French colonization. This is appropriate given the evidence for early modern human behavior and life history in the Maghreb (Bouzouggar et al. 2007; T. Smith et al. 2007), the diversity of various epi-Paleolithic and Neolithic cultures in or near the region associated with climatic changes (Lubell et al. 1984; Rahmani 2003, 2004; Sereno et al. 2008; Sheppard and Lubell 1990), and the interactions with known peoples at later dates (Bennett 1960; Brown 1968; Desanges 1981; Hirschberg 1960; Nebel 2002).
In reviewing data from multiple disciplines, Frigi and colleagues have given the regions populations a multidimensional existence. In providing evidence for the ongoing microevolution in the Maghreb of ancient mtDNA lineages that emerged in Africa and evidence of later gene flow from multiple directions, they have revealed that this region has biological continuity with the deep past as well as change.
Frigi and colleagues may have inadvertently revealed peoples whose ancestors had a level of cultural flexibility in accepting outsiders as mates. As noted, the male and female histories of a population may be different in their sources (Wilkins 2006), although they are now seen as part of a recognizable biocultural entity with a categorically singular identity. How is the emergence of the Amazigh peoples in the geographic range of their homeland to be understood in terms of culture, language, and biology? In some sense the question is about origins, a term that can be confusing because of its various meanings. It can be applied to different aspects of a populationwhich can be disarticulated and can change as a function of time. Ancestry must not be confused with explanation, or gene history with population or culture history. Known ancestors and the ancestors of ones genes are not the same things necessarily (Weiss and Long 2009).
Can a narrative of origins be constructed on the basis of an internal perspective of the dynamics of the human communities of northwestern Africa, considered through time? Or is this region simply an appendage of other places? Sometimes the Amazigh, by use of the term Eurasian in a categorical model of analysis, are placed in a raciotypological model without reference to evolution and their indigenous emergence in Africa. (It can be ventured that this is largely based on nonevolutionary ideas about phenotype, notions of bounded unchanging populations, problematic assumptions about language families, and certain old attitudes and theories about Africa.)
Frigi and colleagues have documented the deep-time biological connections of current Berber speakers to Africa. The migration of Europeans and Asians is also discussed. There has been continuity and change in the population from original settlement. It is important to remember that high levels of gene flow and biocultural assimilation could lead to great biological heterogeneity in a population whose language family or culture does not change. Frigi and colleagues address the idea of the indigenous, although not explicitly, and lay the groundwork for more nuanced future discussions. They suggest a complex biogeographic history not reducible to raciotypological constructs or outdated simplistic theories of colonization and migration. They provide a basis for a rich discussion by acknowledging the interactions of known peoples in the Maghreb and unknown actors of a deeper past. The issue of what is indigenous is seen to be one of definition, turning on what aspect of a population or region is ranked as its defining characteristic, and whether this may change or could have changed over time. The term indigenous unfortunately is connected to a discourse about the West and non-West and sometimes has a negative sensibility and hence may not be the best word, but a discussion of this issue is beyond the scope of this presentation. Of course indigenous is a relative term when the temporal scope of human evolution and history is considered, and it even seems to depend to a degree on what part of the world is under discussion. Europe can serve as a good example. If it is asked who are the indigenous Europeans, there would probably be a request to clarify the time depth, given that modern humans are not native to Europe and arrived there from elsewhere.
(The next question therefore is at what point do they become European and what precisely does this mean: current limb proportions, skin color, genetic variation, language, the presence of Neanderthal DNA?) Does indigenousness require residency back to the upper Paleolithic, the Neolithic, and so on? Is it only a biological phenomenon requiring a drop of Neanderthal blood or a linguistic phenomenon requiring the speaking of Indo-European languages? Or if the question is who were the indigenous inhabitants of northern, southern, western, eastern, or central Europe, the answers would necessarily take on a different tone, based on other information.
Are the Basque speakers the indigenous inhabitants of Europe, if currently spoken language phyla and families are used as population markers, a problematic assumption? Basque predates Indo-European, and there is some indication of some level of biological distinctiveness (Alonso et al. 2005). The fact that historical linguists (e.g., Ehret 2002; Nichols 1997) can reconstruct the existence of culture-linguistic units for a proto-language family (e.g., Adamawa) or phylum (e.g., Niger-Congo), which may have migrated, does not mean that they are suggesting that the people making up such entities connote genetic units or Mendelian breeding populations. It also does not mean that the speakers of such proto-entities had a common molecular or social genealogical origin at foundation, or that the linguists are suggesting this.
Defining origins or indigenous becomes one of perspective. How much Basque ancestry would a European population have to have for the label of indigenous European to apply? If none, why not? (What is the relationship between cultural and biological genealogy?) Can it be assumed that the Basques of today biologically represent those of the past accurately? The post-Paleolithic European assimilation of males from Africa and Asia bearing younger genetic variants is documented (Cruciani et al. 2004, 2007): Are such ancient admixed populations to be viewed as less European or non-European? Are Nordics or the Basques the standard European? Is language, biology, culture, geography, or something else the arbiter of European-ness? In practice, this question seems to be little asked in studies of Europeans: All these groups and nationalities are considered European with little question. Aegean peoples are not presented as hybrids. The linguistic and genetic diversity is not a factor in the designation of indigenous for Europe.
But in the case of Africa there seems to be a problem with diversity for some scholars. The Indo-European language phylum, in the standard evidence-based interpretation, did not originate in the European heartland (Ehret, personal communication, 2010). Most people in Europe today speak Indo-European languagesnow considered as indigenous as Basque. What does it mean for the concept of European if Europes major language phylum did not originate in what is considered Europe proper? How much of the spread of early Indo-European was due to outright settler colonization and how much to language shiftthese are questions that will likely be debated for some time. Are the Finns, Saami, and Hungarians (or their original ancestors)all non-Indo- European-speakingto be considered Europeans? Apparently so. Contrast this with ideas held by some about Berbers as Eurasians who speak a language family that belongs to a phylum whose proto-parent emerged in Africa using standard historical linguistic criteria and whose major history and differentiation occurred in Africa (Ehret 2002; Greenberg 1963; Nichols 1997). In discussions about Europe, geography seems to be enough to define what is indigenous with the exception of the Turks. This contrast deserves review by students of the sociology of knowledge.
The European example is relevant to the discussion of Berbers because of the use of terms by some researchers that imply that Berbers are not an African development, an African people, in their beginning and current state. Calling the Amazigh Eurasian based primarily on skin color without a discussion of process in history, language, evolution, and Y-chromosome variants can easily be seen as problematic when literature about Europe is examined carefully. The possibility of asymmetric gene flow with more Eurasian females being assimilated into the ancient Maghreband their lineages simply differentially surviving in greater frequenciesis addressed in a preliminary fashion by Frigi et al. (2010) and further engages us in the history of social interactions that may influence population biology.
Returning to the Amazigh, the findings and comments of Frigi et al. (2010) on Tunisian Berbers, and Berbers in general, suggest a new way of looking at the Maghreb region of Africa. Their review and analysis offer the opportunity to begin to develop a new and nuanced narrative about the peopling of the region, one that avoids the biases of past writings. Their findings of ongoing evolution in the Maghreb of ancient mtDNA lineages that originated in Africa, synthesized with the evidence of the assimilation of migrants (mainly female?) from Europe and the Near East, the predominance of uniquely African Y-chromosome lineages, and the observation that Berber is the only extant indigenous language in the region suggest the workings of both biological and cultural processes. There are clearly different levels of biological and cultural history. Except in situations of migrationist settler colonialism associated with the annihilation or conquest of local peoples, groups emerge from local elements and new additionsall influenced by the social and physical environments. This view of populations as assemblages and processes is different from a notion of them as essentialist primordial entities with fixed traits having continuity over time. In any geographic space groups can interact at various levels with various strictures; languages can be adopted partly or fully, and social rules may allow the acceptance of offspring by foreign females but not males, or vice versa. It is possible for a group to view itself as genealogically homogeneous by memory but to evince a genetic heterogeneity of lineages obtained in the remote past. Admixture in the late Pleistocene in the deep background of a regional population is to be differentiated from gene flow between known historical entities.
Frigi and colleagues suggestion that supra-Saharan Africans are an indigenous development with a complex story is a corrective to past models. The settlement of the coastal Maghreb in the Middle and Late Stone Age is a part of the settlement of the world outside Saharo-tropical Africa. The early modern human presence in the Maghreb suggests that that region played a role in modern human developments (Bouzouggar et al. 2007). Even if whole communities later came from outside Africa into the Maghreb (before the Phoenicians), which is not knowable, they became thoroughly assimilated into the autochthonous population which adopted some of its culture (Camps 1982), and their descendants are a part of the emergence of the much later Amazigh world. Less arid climatic conditions in the early to mid-Holocene Sahara allowed for the interaction of various peoples who no doubt contributed to the population history, as observed by Frigi et al. (2010). Saharan developments likely help to explain the Berber emergence, because, based on recent work [see Kuper and Kropelin (2006) and Sereno et al. (2008)], the desert was likely the site of a metapopulation and cultural differentiation. Whether the early Saharan rock paintings depict only Africans of varying phenotypes or such Africans and Asians (as suggested by Frigi and colleagues) can be debated, but the net result was assimilation into Amazigh communities, because there are no Berbers in Europe or Asia. The light skin color of Mediterranean Africa may be the result of adaptive evolution or drift, given the length of time of modern people in Africa, including the Maghreb, or gene flow, but more likely some combination.
The Maghreb has several Neolithic traditions (Camps 1982; Phillipson 2005), which might indicate different peoples or simply cultural adoption or adaptation by heterogeneous populations who became unified under singular cultural practices and one language family. The Neolithic Capsian tradition shows continuity with previous cultures, with evidence of these accepting domesticated sheep and goat into a local subsistence pattern, thus becoming Neolithicized with a pastoralist economy (Rahmani, 2003, 2004; Sheppard and Lubell 1990). A. B. Smith (2005) and McDonald (1998) indicate the importance of pastoralism in the Holocene Sahara, and this economy may help in the understanding of Berber emergence. In the coastal Maghreb various Neolithic and post-Neolithic interregional interactions are in evidence, based on archaeology and the eventual settlements of the Phoenicians, Romans, Vandals, and others (Camps 1982).
In aggregate, over time, these peoples, along with the later importation of Europeans (Bennett 1960; Davis 2004), would seemingly have contributed far more to the current biological picture than has been realized. The much later trans-Saharan trade in enslaved individuals no doubt played a role in genetic contributions, but the egress from a desiccating Sahara with subsequent population formations would explain some of the younger sub-Saharan variation, be it from western or eastern Africa. The Eurasian component seems to have come in over a longer period of time, as noted earlier. A small amount of gene flow per generation into a population or geographic region can drastically change its original gene frequencies in only a few thousand years, as noted by Cavalli-Sforza (1991).
The development of a narrative of the population history of the Maghreb requires careful analysis using several approaches. Frigi and colleagues have made an important contribution to studies of African human biology and culture in suggesting the complexity of Maghreban population history."
Recent DNA study shows Berber roots foundational in africa and African populations with deep-rooted African DNA lineages not merely migration from the outside/
"Our objective is to highlight the age of sub-Saharan gene flows in North Africa and particularly in Tunisia... The results show that the most ancient haplogroup is L3*, which would have been introduced to North Africa from eastern sub-Saharan populations around 20,000 years ago. Our results also point to a less ancient western sub-Saharan gene flow to Tunisia, including haplogroups L2a and L3b. This conclusion points to an ancient African gene flow to Tunisia before 20,000 years BP. These findings parallel the more recent findings of both archaeology and linguistics on the prehistory of Africa. The present work suggests that sub-Saharan contributions to North Africa have experienced several complex population processes after the occupation of the region by anatomically modern humans. Our results reveal that Berber speakers have a foundational biogeographic root in Africa and that deep African lineages have continued to evolve in supra-Saharan Africa."
-- Ancient Local Evolution of African mtDNA Haplogroups in Tunisian Berber Populations Frigi et al. Human Biology (August 2010 (82:4)
"Our findings are in accordance with other studies on Y-chromosome markers that have shown that the predominant Y-chromosome lineage in Berber communities is the subhaplogroup E1b1b1b (E-M81), which emerged in Africa, is specific to North African populations, and is almost absent in Europe, except in Iberia (Spain and Portugal) and Sicily. Molecular studies on the Y chromosome in North Africa are interpreted as indicating that the southern part of Africa, namely, the Horn/East Africa, was a major source of population in the Nile Valley and northwest Africa after the Last Glacial Maximum, with some migration into the Near East and southern Europe (Bosch et al. 2001; Underhill et al. 2001).
--Ancient Local Evolution of African mtDNA Haplogroups in Tunisian Berber Populations Frigi et al., 2010
Posts: 5905 | From: The Hammer | Registered: Aug 2008
| IP: Logged |
I hope you can comprehend this quote from your cut and paste:
the Muurs and the Wild-mans :
quote: n any case, our results show that Neandertal genomic traces do not mark a division between African and non-Africans but rather a division between Sub-Saharan Africans and the rest of modern human groups, including those from North Africa.
Posts: 7419 | From: North America | Registered: Mar 2009
| IP: Logged |
I hope you can comprehend this quote from your cut and paste:
the Muurs and the Wild-mans :
quote: n any case, our results show that Neandertal genomic traces do not mark a division between African and non-Africans but rather a division between Sub-Saharan Africans and the rest of modern human groups, including those from North Africa.
Dufus, that statement is only reiteretaing what the title of the study tells you
posted
When Neanderthal admixture in Noth Africans is mentioned zarahan pulls out articles that suggest humans may not have mixed with Neanderthals.
When Europeans are mentioned he posts articles which say that are mixed with Neanderthals.
quote:Originally posted by zarahan- aka Enrique Cardova:
Pure white genetics produced only the savage Neanderthals. Africa by contrast, produced a better, more advanced human than white lineages. The first pure white genetic output was thus brutish, primitive, dead end Neanderthals. By contrast, it was Africa that produced the future of humanity- anatomically modern humans, not cold-climate Europe. White NEanderthals survived, until they were supplanted by more advanced, modern tropically adapted humans from Africa that mingled with and replaced them. This would be the first instance of the hybridization that produced today's white Europeans. It was the coming of tropically adapted African variants into Europe that caused Europe to progress beyond the brutish pure white Neanderthal lineages.
.
that's why he's a black human biodiversity proponent when it's convenient when it's not convenient he's a sweet lil angel In other words a clear racist and hypocrite
Posts: 42940 | From: , | Registered: Jan 2010
| IP: Logged |
I hope you can comprehend this quote from your cut and paste:
the Muurs and the Wild-mans :
quote: n any case, our results show that Neandertal genomic traces do not mark a division between African and non-Africans but rather a division between Sub-Saharan Africans and the rest of modern human groups, including those from North Africa.
Dufus, that statement is only reiteretaing what the title of the study tells you
Duncey
The man in the picture is a mulatto. Part of his ancestry came from central Asia via the Arabian Gulf.
Besides, I understand from Van Sertima Vindicated that Neanderthal is a 48 chromosomal monkey.
posted
My view is if you pay taxes to monkeys, if you go to the hospital you are treated by monkey doctors, if you pay monkeys a monthly internet bill, if monkeys have the most advanced mlitary, if you were educated in a school run by monkeys, if you pay monkeys rent, then it's hard to argue the monkey has a lower IQ. I mean look at reality for a moment
Posts: 42940 | From: , | Registered: Jan 2010
| IP: Logged |
posted
hen Europeans are mentioned he posts articles which say that are mixed with Neanderthals.
The article posted questioned Neanderthal admixture across the board- for all humans, not just "when Europeans are mentioned."
And, you conveniently, as you usually do, forget to mention the full context of my quote. Now why is that liaress? I clearly target the hypocritical "biodiversity" racial approach and say, that if these methods are used, then certain results follow. Its you who are the racist and hypocrite.
full context which you "conveniently" leave out- quote:
"Some white "Racial Reality" or "biodiversity" proponents argue for European descent from NEanderthals "untainted" by Africa- a lily white "pure" line of white evolution... let's go with their "pure white" evolution approach for a moment."
Whatsa matta? When I apply your racist "biodiversity" approach all of a sudden you are offended? I mean you are all for a picture of virtuously evolved white people untainted by "the negroes," but when I take your own racist approach and flip it to apply its full "racial" implications where all of a sudden your white "role models" are not in such a good light, you can't stand the heat?
-------------------- Note: I am not an "Egyptologist" as claimed by some still bitter, defeated, trolls creating fake profiles and posts elsewhere. Hapless losers, you still fail. My output of hard data debunking racist nonsense has actually INCREASED since you began.. Posts: 5905 | From: The Hammer | Registered: Aug 2008
| IP: Logged |
quote:Originally posted by zarahan- aka Enrique Cardova: hen Europeans are mentioned he posts articles which say that are mixed with Neanderthals.
The article posted questioned Neanderthal admixture across the board- for all humans, not just "when Europeans are mentioned."
And, you conveniently, as you usually do, forget to mention the full context of my quote. Now why is that liaress? I clearly target the hypocritical "biodiversity" racial approach and say, that if these methods are used, then certain results follow. Its you who are the racist and hypocrite.
full context which you "conveniently" leave out- quote:
"Some white "Racial Reality" or "biodiversity" proponents argue for European descent from NEanderthals "untainted" by Africa- a lily white "pure" line of white evolution... let's go with their "pure white" evolution approach for a moment."
Whatsa matta? When I apply your racist "biodiversity" approach all of a sudden you are offended? I mean you are all for a picture of virtuously evolved white people untainted by "the negroes," but when I take your own racist approach and flip it to apply its full "racial" implications where all of a sudden your white "role models" are not in such a good light, you can't stand the heat?
you are a proponent of the Iceman Inheritance, stop fronting
My mans Clyde and Marc say the neanderthal was black anyway.
Here's what happened, way back before 30,000 years ago when there were no white peoples, primitive man raped Neanderthals. Eventually he killed them all off. Then he said they were savages and deserved it. But the neanderthals would tell a different story > genocide
Posts: 42940 | From: , | Registered: Jan 2010
| IP: Logged |
This lecture was delivered by Dr. Ian Tattersall at The Metropolitan Museum of Art on the occasion of the symposium "Genesis: Exploration of Origins" on March 7, 2003. This symposium was held in conjunction with the special exhibition, "Genesis: Ideas of Origin in African Sculpture," and was made possible through the support of The Ford Foundation.
quote:At about 40,000 years ago, however, Homo sapiens, in the form of the Cro-Magnons, began trickling into Europe, probably from an initially African place of origin.
[...]
It was brought with them by the Cro-Magnons, whose new qualities had emerged elsewhere. Probably this was in Africa, for it is from this continent that we have not just the first suggestions of the emergence of modern anatomical structure, but of modern behaviors as well.
[...]
The most remarkable early evidence of symbolic activity in Africa comes in the form of the recent find of engraved ochre plaques, such as this one, from Blombos Cave on the southern coast of Africa (Fig. 10). This is an unequivocally symbolic object, even if we cannot directly discern the significance of the geometric design that the plaque bears; and it is dated to around 70,000 years ago, over 30,000 years before anything equivalent is found in Europe.
To evidence such as this can be added suggestions of a symbolic organization of space at the site of Klasies River Mouth (Fig. 11), also near the southern tip of Africa, at over 100,000 years ago. Pierced shells, with the strong implication of stringing for body ornamentation, are known from Porc-Epic Cave in Ethiopia at around 70,000 years ago. Bone tools of the kind introduced much later to Europe by the Cro-Magnons, are found at the Congolese site of Katanda, dated to perhaps 80,000 years ago. Blade tool industries, again formerly associated principally with the Cro-Magnons, are found at least sporadically at sites in Africa that date to as much as a quarter of a million years ago. Also in the economic/technological realm, such activities as flint-mining, pigment-processing and long-distance trade in useful materials are documented in Africa up to about 100,000 years ago. These and other early African innovations are reviewed by McBrearty and Brooks (2000).
quote:Originally posted by the lioness,: I'm approaching 17.000 posts now. I forgot I even made this thread
17.000 posts of euronut B.S.
-- Anwar G. Chejne Islam and the West: The Moriscos, a Cultural and Social History(1993)
Posts: 22244 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |





 UBBFriend: Email this page to someone!
UBBFriend: Email this page to someone!









![[Big Grin]](biggrin.gif) :
:





 Printer-friendly view of this topic
Printer-friendly view of this topic