Y chromosome evidence of earliest modern human settlement in East Asia and multiple origins of Tibetan and Japanese populations 2013
Hong Shi1, Hua Zhong2, Yi Peng1, Yong-Li Dong3, Xue-Bin Qi1, Feng Zhang4, Lu-Fang Liu5, Si-Jie Tan3, Runlin Z Ma2, Chun-Jie Xiao3, R Spencer Wells6, Li Jin4 and Bing Su1*
Abstract Background The phylogeography of the Y chromosome in Asia previously suggested that modern humans of African origin initially settled in mainland southern East Asia, and about 25,00030,000 years ago, migrated northward, spreading throughout East Asia. However, the fragmented distribution of one East Asian specific Y chromosome lineage (D-M174), which is found at high frequencies only in Tibet, Japan and the Andaman Islands, is inconsistent with this scenario.
Results In this study, we collected more than 5,000 male samples from 73 East Asian populations and reconstructed the phylogeography of the D-M174 lineage. Our results suggest that D-M174 represents an extremely ancient lineage of modern humans in East Asia, and a deep divergence was observed between northern and southern populations.
Conclusion We proposed that D-M174 has a southern origin and its northward expansion occurred about 60,000 years ago, predating the northward migration of other major East Asian lineages. The Neolithic expansion of Han culture and the last glacial maximum are likely the key factors leading to the current relic distribution of D-M174 in East Asia. The Tibetan and Japanese populations are the admixture of two ancient populations represented by two major East Asian specific Y chromosome lineages, the O and D haplogroups.
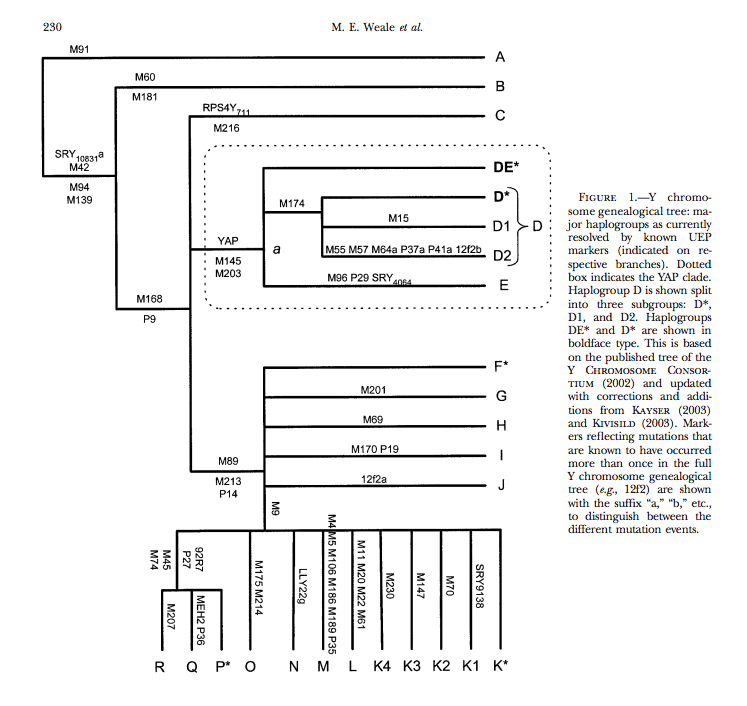
Introduction The Y chromosome Alu polymorphism (YAP, also called M1) defines the deep-rooted haplogroup D/E of the global Y-chromosome phylogeny [1]. This D/E haplogroup is further branched into three sub-haplogroups DE*, D and E (Figure 1). The distribution of the D/E haplogroup is highly regional, and the three subgroups are geographically restricted to certain areas, therefore informative in tracing human prehistory (Table 1). The sub-haplogroup DE*, presumably the most ancient lineage of the D/E haplogroup was only found in Africans from Nigeria [2], supporting the "Out of Africa" hypothesis about modern human origin. The sub-haplogroup E (E-M40), defined by M40/SRY4064 and M96, was also suggested originated in Africa [3-6], and later dispersed to Middle East and Europe about 20,000 years ago [3,4]. Interestingly, the sub-haplogroup D defined by M174 (D-M174) is East Asian specific with abundant appearance in Tibetan and Japanese (3040%), but rare in most of other East Asian populations and populations from regions bordering East Asia (Central Asia, North Asia and Middle East) (usually less than 5%) [5-7]. Under D-M174, Japanese belongs to a separate sub-lineage defined by several mutations (e.g. M55, M57 and M64 etc.), which is different from those in Tibetans implicating relatively deep divergence between them [1]. The fragmented distribution of D-M174 in East Asia seems not consistent with the pattern of other East Asian specific lineages, i.e. O3-M122, O1-M119 and O2-M95 under haplogroup O [8,9].
Besides Tibetans and Japanese, D-M174 is also prevalent in several southern ethnic populations in East Asia, including the Tibeto-Burman speaking populations from Yunnan province of southwestern China (14.072.3%), one Hmong-Mien population from Guangxi of southern China (30%) and one Daic population from Thailand (10%), which could be explained by fairly recent population admixture [9-11]. However, a recent study reported a high frequency of D-M174 in Andamanese (56.25%), people who live in the remote islands in the Indian Ocean and considered one of the earliest modern human settlers of African origin in Southeast Asia [12]. Another study by Underhill et al. suggested that the D-M174 lineage likely reached East Asia about 50,000 years ago [5]. This implies that the YAP lineage in East Asia could be indeed very ancient.
The distribution patterns of the four D-M174 sub-lineages (sub-haplogroups) (Figure 1) are different from each other. D1-M15 is widely distributed across East Asia including most of the Tibeto-Burman and Daic speaking populations (Table 2). D*-M174 and D3-p47 are mainly distributed in Tibeto-Burman populations with sporadic occurrence in the Daic populations. In surprise, we observed two DE* in the Tibetan samples, which was previously only observed in Africa (Nigerians), but not in other world populations. In contrast, D2-M57 only occurred in Japanese, an implication of the early divergence of this lineage from other D-M174 sub-haplogroups (Table 2). We identified four E-M40 individuals in the northwestern Han populations, a reflection of recent gene flow from Central Asia.
The East African megadroughts (about 13575 thousand years ago) during the early late-Pleistocene was suggested compelling modern humans out of Africa [31]. And the early modern human could occupy coastal areas and exploited the near-shore marine food resource by that time [32]. Then, modern humans was suggested expanding along the tropical coast, and the earliest modern human fossil found out of Africa was about 100,000 years ago [33]. The period of 80,00010,000 years ago during the last glacial might have a huge impact on modern human migration, and the sea level had fallen 50200 meters below present [34], which resulted in larger dry lands and possibility for human migration between currently separated lands by ocean, e.g. between Japan and the mainland.
The divergence time of haplogroup D is about 60,000 years ago, considering the wide though fragmented geographic distribution of D-M174, the proposed Paleolithic migration would be the first northward population movement of modern humans after their initial settlement in southern East Asia. As the last glacial occurred during 80,00010,000 years ago, the northward migration of D-M174 is consistent with the proposed notion that modern humans might exploit the food of "Mammoth Steppe" [39]. Besides the later population expansion, the cold weather during the last glacial may also contribute to the current fragmented distribution of D-M174. Interestingly, a recent archaeological finding supported that modern humans explored the Tibetan plateau about 30,00040,000 years ago, which is much earlier than previously suggested [40], but consistent with our hypothesis. The after-glacial sea level rise eventually led to the separation between Japan and the main continent, which explains the relic distribution of D-M174 in current Japanese populations. The archaeological data suggested that the initial colonization of modern humans in Japan occurred about 30,000 years ago [41,42], consistent with our age estimation of D2-M57 (37,678 ± 2,216 years ago). Taken together, the current Tibetan and Japanese populations are probably the admixture of two ancient populations represented by D-M174 and O3-M122 respectively [7,10,16].
Conclusion In summary, we demonstrated an ancient Paleolithic population migration in East Asia, predating the previously suggested northward population movement. The current fragmented distribution of D-M174 is likely due to the combination of later Neolithic population expansion and the last glacial.
Posts: 42940 | From: , | Registered: Jan 2010
| IP: Logged |
posted
The do not mention another possibility- that the divergences actually BEGAN INSIDE Africa via the "root types" that yielded "E". There could still be "back-migration" of the proto "E" groups, in which case, they are migrating back to the point of origin where root types of "E" first began to emerge. There is some indication that this is what happened with various mtDNA sequences. Like the mtDNA, the divergence could have taken place somewhere between a point of origin in Africa and say, India. Yan et al present minimal evidence showing an exclusive development outside of Africa.
"These indicate that the root of L3 gives rise to a multifurcation from a single haplotype producing a number of distinct subclades... The simplest explanation for this geographical distribution [haplogroups M and N], however, is an expansion of the root type within East Africa, where several independent L3 branches thrive, including a sister group to L3, christened L4 (Kivisild et al. 2004; Chap. 7), followed by divergence into haplogroups M and N somewhere between the Horn of Africa and the Indian subcontinent. Since neither the L3 root type nor any other descendants survive outside Africa, the root type itself must have become extinct during a period of genetic drift in the founder population as it diversified into haplogroups M and N, if the diversification was outside Africa. If on the other hand the diversification was indeed within East Africa, then Haplogroups M and N must have either been carried out of Africa in their entirety or subsequently have become extinct within Africa, with the singular exception of the derived M1." - Hans-Jürgen Bandelt et. 2006. EDS. Human Mitochondrial DNA and the Evolution of Homo sapiens.
And as Keita notes too often claims of "Eurasian" origin can be misleading as regards African peoples.
In any event, since 75% of all Y-chromosomes found in Africa belong to "E", even if any ancient tribes moved into the continent from say Sinai or other points, they are still African, and looked like today's Africans.
-------------------- Note: I am not an "Egyptologist" as claimed by some still bitter, defeated, trolls creating fake profiles and posts elsewhere. Hapless losers, you still fail. My output of hard data debunking racist nonsense has actually INCREASED since you began.. Posts: 5905 | From: The Hammer | Registered: Aug 2008
| IP: Logged |
quote:Originally posted by xyyman: I did not read the paper as yet ...but...are modern day Africans...Asians?....hmmm!!
quote:Originally posted by the lioness,: Asiacentricity?
the PDF link is at the top of the thread
quote:Originally posted by zarahan- aka Enrique Cardova:
In any event, since 75% of all Y-chromosomes found in Africa belong to "E", even if any ancient tribes moved into the continent from say Sinai or other points, they are still African, and looked like today's Africans.
the second article which is a reference (second post) of the first article speaks of hap DE:
"The sub-haplogroup DE*, presumably the most ancient lineage of the D/E haplogroup was only found in Africans from Nigeria"
Y chromosome evidence of earliest modern human settlement in East Asia and multiple origins of Tibetan and Japanese populations 2013
Hong Shi1,
_________________________
The majority of DE male lines can be categorized as being in either Haplogroup D (Y-DNA), which likely originated in Asia, the only place where it has been found,[1] or haplogroup E, which is believed to have originated in East Africa[3][4] or the Near East.[5] The remainder are said to be in the paragroup DE*, confirmed cases of which are extremely rare.
wiki:
Haplogroup DE
In a study of over 8000 men worldwide including 1247 Nigerians, Haplogroup DE* was observed in only 5 Nigerian males (5/1247). However, the study's authors caution that "the apparently paraphyletic status of this haplogroup, and hence the conclusions of nested cladistic analysis, are also likely to be illusory" and that "the only genealogically meaningful definition of the age of a clade is the time to its most recent common ancestor, but only if DE* is paraphyletic does it also become automatically older than D or E in this sense."[6] More recently, one example of DE* was found amongst the Nalu in Guinea Bissau (1/17). The DE* sequence of this individual differs by one mutation from the DE* sequence of the Nigerian individuals. This indicates common ancestry, though the phylogenetic relationship between the two lineages was not determined in this particular study.[7] A 2008 study detected DE* in two individuals from Tibet (2/594).[8]
Haplogroup DE is found in Africa (Haplogroups E and DE*) and East Asia (Haplogroups D and DE* and E*) but is largely absent in between these two regions. The presence of DE across widely separated regions has confounded investigators trying to reconstruct the migration of humans from Africa to Asia. At some time, there was an extinction of DE lineages in West, South and Central Asia. Autochthonous DE lineages are absent in India, an important region in the dispersal of humans in Asia. However DE lineages have been detected in relict populations of the Andaman Islands. Underhill et al. 2007, suggest the possibility that deleterious mutations in some DE carriers may explain the extinction of DE lineages in India.[9]
The YAP insertion was discovered by scientists led by Michael Hammer of the University of Arizona.[10] Between 1997 and 1998 Hammer published three articles relating to the origins of haplogroup DE.[11][12][13] These articles state that YAP insertion occurred in Asia. As recently as 2007, some studies such as Chandrasekar et al. 2007, cite the publications by Hammer when arguing for an Asian origin of the YAP insertion.[5]
The scenarios outlined by Hammer include an out of Africa migration over 100,000 years ago, the YAP+ insertion on an Asian Y-chromosome 55,000 years ago and a back migration of YAP+ from Asia to Africa 31,000 years ago by its subclade haplogroup E.[13] This analysis was based on the fact that older African lineages, such as haplogroups A and B, were YAP negative whereas the younger lineage, haplogroup E was YAP positive. Haplogroup D, which is YAP positive, was clearly an Asian lineage, being found only in East Asia with high frequencies in Japan and Tibet. Because the mutations that define haplogroup E were observed to be in the ancestral state in haplogroup D, and haplogroup D at 55kya, was considerably older than haplogroup E at 31kya, Hammer concluded that haplogroup E was a subclade of haplogroup D
Contemporary studies[edit] In 2000 a number of scientists had started to reassess the hypothesis of an Asian origin of the YAP insertion. Underhill et al. 2000 identified the D-M174 mutation that defines haplogroup D. The M174 allele is found in the ancestral state in all African lineages including haplogroup E. The discovery of M174 mutation meant that haplogroup E could not be a subclade of haplogroup D. These findings effectively neutralized the argument of an Asian origin of the YAP+ based on the character state of the M40 and M96 mutations that define haplogroup E. According to Underhill et al. 2000, the M174 data alone would support an African origin of the YAP insertion.[14]
Further arguments were made supporting and African origin of the YAP in Underhill et al. 2001. The arguments for an African origin include.[3]
Africa has the highest frequency of YAP(>80%). Whereas the YAP+ in Asia has a fairly restricted geographic distribution, mainly at low to moderate frequencies (average 9.6%) in East Asia.[8] It was claimed that there was no archaeological evidence of a back-migration to Africa, and at the time of writing that there was no unequivocal Y DNA, mitochondrial DNA or autosomal DNA evidence of a back migration to Africa.[3] Although Haplogroup C seems to have originated in Asia at a similar time to Haplogroup DE's origin, Haplogroup C shows no sign of back migration to Africa. The African origin of the YAP+ is also supported by studies concerning haplogroup E. In Altheide and Hammer 1997, the authors argue that haplogroup E arose in Asia on an ancestral YAP+ allele before migrating back to Africa.[12] However some studies, such as Semino et al., indicate that the highest frequency and diversity of haplogroup E is in Africa, and Africa is the most likely place of origin of the haplogroup.[8][15]
The models supporting an African origin or an Asian origin of the YAP+ insertion both required the extinction of the ancestral YAP chromosome to explain the current distribution of the YAP+ polymorphism. Paragroup DE* possesses neither the mutations that define haplogroup D or haplogroup E. If paragroup DE* was found in one location but not the other, it would boost one theory of the other.[16] Haplogroup DE* has recently been found in Nigeria,[6] Guinea-Bissau[7] and also in Tibet.[8] The phylogenetic relationship of three DE* sequences has yet to be determined, but it is known that the Guinea Bissau sequences differ from the Nigerian sequences by at least one mutation.[7] Weale et al. state that the discovery of DE* among Nigerians pushes back the date for the most recent common ancestor (MRCA) of African YAP chromosomes. This, in his view, has the effect of reducing the time window through which a possible back migration from Asia to Africa could occur.[6]
Chandrasekhar et al. 2007, have argued for the Asian origin of the YAP+. They state,
The presence of the YAP insertion in Northeast Indian tribes and Andaman Islanders with haplogroup D suggests that some of the M168 chromosomes gave rise to the YAP insertion and M174 mutation in South Asia
They also argue that YAP+ migrated back to Africa with other Eurasian haplogroups. These include Haplogroup R1b1* (18-23kya), which has been observed with especially high frequency among the members of some peoples in northern Cameroon, and Haplogroup T (25-30kya), which has been observed in low frequencies in Africa. Haplogroup E at 50kya is considerably older than these haplogroups and has been observed at frequencies of 80-92% in Africa.
In a press release concerning a study by Karafet et al. (2008), Michael Hammer, revised the dates for the origin Haplogroup DE from 55,000 years ago to 65,000 years ago. For haplogroup E, Hammer revised the dates from 31,000 years ago to 50,000 years ago. Hammer is also quoted as saying The age of haplogroup DE is about 65,000 years, just a bit younger than the other major lineage to leave Africa, which is assumed to be about 70,000 years old, in which he implies that haplogroup DE left Africa along with Haplogroup CF.[17]
Peter Underhill states that there will always be uncertainty regarding the precise origins of DNA sequence variants such as YAP because of a lack of knowledge concerning prehistoric demographics and population movements. However Underhill contends that with all the available information, the African origin of the YAP+ polymorphism is more parsimonious and more plausible than the Asian origin hypothesis.[9] Other authors who have published or co-published works in support of an African origin of the YAP+ include Luigi Luca Cavalli-Sforza,[14] Toomas Kivisild,[9] Spencer Wells,[16] Linda Stone and Paul F. Lurquin.[18]
Posts: 42940 | From: , | Registered: Jan 2010
| IP: Logged |
posted
And the people in Europe are not really Europeans.........
-------------------- Without data you are just another person with an opinion - Deming Posts: 12143 | From: When you have eliminated the impossible, whatever remains, however improbable | Registered: Jun 2007
| IP: Logged |
posted
The Canadian criminal justice system should move away from the idea that DNA evidence stands on its own to prove guilt, according to former Supreme Court justice Ian Binnie.
DNA has taken on this magic and there are so many wrongful convictions that have not only been exposed by DNA but which have been inflicted by DNA, said Binnie during a discussion panel on the evidence revolution in Canada last week at the Canadian Institute for the Administration of Justices annual conference held in Toronto.
The reliance on genetic proof ignores many possibilities that would make a piece of evidence questionable, said Binnie, who gave the example of a mix-up of samples at a laboratory.
Canada should follow Australias lead by introducing a provision that says DNA proof is conclusive only in the presence of other corroborative evidence pointing to the same truth, according to Binnie....
quote:Originally posted by xyyman: I did not read the paper as yet ...but...are modern day Africans...Asians?....hmmm!!
quote:Originally posted by the lioness,: Asiacentricity?
It's an interesting story/ theory, the source references material they used is rather relatively old. The youngest sources dates back to 2010.
quote: According to the phylogenetic tree of Y chromosome, all the modern males could be categorized into 20 major monophyletic or paraphyletic groups (referred to as A to T) and their subclades 13,14. Nearly all the Y chromosomes outside Africa are derivative at the SNP M168 and belong to any of its three descendent super-haplogroups DE, C, and F 9,10,15, strongly supporting the out-of-Africa theory. The time of the anatomically modern humans exodus from Africa has yielded inconsistent results ranging from 39 kya 16, 44 kya 10, 59 kya 17, 68.5 kya 18 to 57.0 74.6 kya 19.
To achieve sufficiently high coverage in the non-recombining regions of Y chromosome (NRY) and an adequate representation of individual samples, we selected 110 males, encompassing the haplogroups O, C, D, N, and Q which are common in East Eurasians, as well as haplogroups J, G, and R which are common in West Eurasians (see Table S1), and sequenced their non-repetitive segments of NRY using a pooling-and-capturing strategy.
Results
Overall ~4,500 base substitutions were identified in all the samples from the whole Y chromosome, in which >4,300 SNPs that has not been publicly named before 2012 (ISOGG etc.). We designated each of these SNP a name beginning with F (for Fudan University) (see Table S2).
[...]
The first divergence event out of Africa, i.e. between Haplogroup DE and the ancestor of C and F, is dated at 54.1 kya (95%CI 50.6 58.2), inside the range of previous estimations. Within the 3.9 Mbp range, only 3 SNPs were observed between the divergence events of DE/CF and C/F, indicating that DE, C, and F likely emerged subsequently in less than a thousand years. After diverged from Haplogroup C, no major split was observed in F for 18 thousand years, suggesting a strong bottleneck of F lineage. It should be noted that all the primary haplogroups (G, J, N, O, Q, and R) emerged before the last glacial maximum (LGM, ~20 kya), and most of the presently known East Eurasian clades have branched off in the late Upper Paleolithic Age (before 10 kya). Only binary divergences on this tree occurred before 7 kya, suggesting that during the Paleolithic Age, slow population growth and frequent bottlenecks eradicated most of the ever existing clades 22.
--Shi Yan (2013)
Y Chromosomes of 40% Chinese Are Descendants of Three Neolithic Super-grandfathers
quote: The DE haplogroup appeared approximately 50,000 years bp in North East Africa and subsequently split into haplogroup E that spread to Europe and Africa and haplogroup D that rapidly spread along the coastline of India and Asia to North Asia.
quote: A1a-M31 is observed in northwestern Africans; A1b1a-M14 is seen among click language-speaking Khoisan populations. A1b1b-M32 has a wide distribution including Khoisan speaking and East African populations, and scattered members on the Arabian Peninsula
--Haplogroups CT and CF: Everyone who doesn't belong to Haplogroup A or B belongs to the super-group CT, which is defined by mutations M168 and M294. This is the branch of humanity that first left Africa, although the mutation that led to CT probably happened in Africa about 50,000 - 60,000 years ago, in a man from East Africa, who's been called the "Eurasian Adam", since his descendents include all Eurasian and Asian people today. Haplogroup CT was probably the group involved in the early migration out of Africa, which spread along the southern coast of the Arabian peninsula, Iran, Pakistan, India, and all the way to southeast Asia. Here are some maps showing these early migrations. As you can see, the dates calculated from the genetic evidence correspond only roughly to dates determined by the archeological evidence. These are not yet exact sciences in terms of dating, so we have to reckon with some degree of uncertainty. Haplogroup CF diverged soon after, also in Africa, and is defined by mutation P143. Both super-groups CT and CF are found all over the world, including the Americas.
--Interesting, so this should means that a of YAP occurred splitting this CT into C and T, causing mutations in the loci, just like the DE*, as mankind left the East African coast and moved into another region.
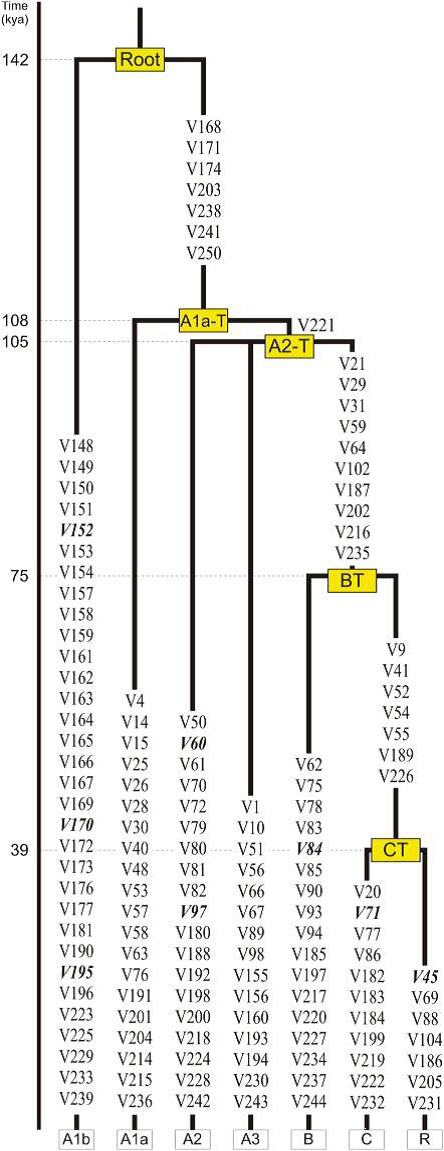
Y-DNA haplogroup A contains lineages deriving from the earliest branching in the human Y chromosome tree. The oldest branching event, separating A0-P305 and A1-V161, is thought to have occurred about 140,000 years ago. Haplogroups A0-P305, A1a-M31 and A1b1a-M14 are restricted to Africa and A1b1b-M32 is nearly restricted to Africa. The haplogroup that would be named A1b2 is composed of haplogroups B through T. The internal branching of haplogroup A1-V161 into A1a-M31, A1b1, and BT (A1b2) may have occurred about 110,000 years ago. A0-P305 is found at low frequency in Central and West Africa. A1a-M31 is observed in northwestern Africans; A1b1a-M14 is seen among click language-speaking Khoisan populations. A1b1b-M32 has a wide distribution including Khoisan speaking and East African populations, and scattered members on the Arabian Peninsula.
quote: Y-DNA haplogroup B, like Y-DNA haplogroup A, is seen only in Africa and is scattered widely, but thinly across the continent. B is thought to have arisen approximately 50,000 years ago. These haplogroups have higher frequencies among hunter-gather groups in Ethiopia and Sudan, and are also seen among click language-speaking populations. The patchy, widespread distribution of these haplogroups may mean that they are remnants of ancient lineages that once had a much wider range but have been largely displaced by more recent population events.
Some geographic structuring is seen between the sub-groups B2a (B-M150) and B2b (B-M112). Sub-group B2b is seen among Central African Pygmies and South African Khoisan. Sub-group B2a is seen among Cameroonians, East Africans, and among South African Bantu speakers. B2a1a (B-M109) is the most commonly seen sub-group of B2a. About 2.3% of African-Americans belong to haplogroup B - with 1.5% of them belonging to the sub-group B2a1a.
quote: Y-DNA haplogroup F is the parent of all Y-DNA haplogroups G through T and contains more than 90% of the worlds population. Haplogroup F was in the original migration out of Africa, or else it was founded soon afterward, because F and its sub-haplogroups are primarily found outside, with very few inside, sub-Saharan Africa. The founder of F could have lived between 60,000 and 80,000 years ago, depending on the time of the out-of-Africa migration.
The major sub-groups of Haplogroup F are Haplogroups G, H, [IJ], and K, which are discussed elsewhere at this site. The minor sub-groups, F*, F1, and F2 have not been well studied, but apparently occur only infrequently and primarily in the Indian subcontinent. F* has been observed in two individuals in Portugal, possibly representing a remnant of 15th and 16th century contact of Portugal with India.
quote: The deepest branching separates A1b from a monophyletic clade whose members (A1a, A2, A3, B, C, and R) all share seven mutually reinforcing derived mutations (five transitions and two transversions, all at non-CpG sites). To retain the information from the reference MSY tree13 as much as possible, we named this clade A1a-T (Figure 1). Within A1a-T, the transversion V221 separates A1a from a monophyletic clade (called A2-T) consisting of three branches: A2, A3, and BT, the latter being supported by ten mutations (Figure 1).
SNP Location Haplogroup Mutations M5 M C > T M9 K, KR C > G M11 L A > G M45 P, PR G > A M69 H T > C M89 F, FR C > T M96 E G > C M122 O3 T > C M168 CR C > T M170 I A > C M174 D T > C M175 O T > A M20 G G > T M207 R A > G M214 NO T > C M304 J A > C M343 R1b C > A P36 Q G > T SRY10831.1 BR A > G
quote:Y-DNA haplogroup A contains lineages deriving from the earliest branching in the human Y chromosome tree. The oldest branching event, separating A0-P305 and A1-V161, is thought to have occurred about 140,000 years ago. Haplogroups A0-P305, A1a-M31 and A1b1a-M14 are restricted to Africa and A1b1b-M32 is nearly restricted to Africa. The haplogroup that would be named A1b2 is composed of haplogroups B through T. The internal branching of haplogroup A1-V161 into A1a-M31, A1b1, and BT (A1b2) may have occurred about 110,000 years ago. A0-P305 is found at low frequency in Central and West Africa. A1a-M31 is observed in northwestern Africans; A1b1a-M14 is seen among click language-speaking Khoisan populations. A1b1b-M32 has a wide distribution including Khoisan speaking and East African populations, and scattered members on the Arabian Peninsula.
Haplogroup CT (M168): Time of Emergence: 70,000 BP, 2800 generations ago beginning of the Last Glacial Period Place of Origin: The African Rift Valleymore by Gábor Balogh
TP's method of argumention is to swamp the issue with mega posts. You start a thread and he starts a new thread within your thread at the same time (he's trying to compete with zarahan) Like if this was a debate at a university one guy would would have a 15 minute opening statement and then Troll would come on and at 2AM he he'd still be there on the opening statement speaking to an empty auditorium
Posts: 42940 | From: , | Registered: Jan 2010
| IP: Logged |
TP's method of argumention is to swamp the issue with mega posts. You start a thread and he starts a new thread within your thread at the same time (he's trying to compete with zarahan) Like if this was a debate at a university one guy would would have a 15 minute opening statement and then Troll would come on and at 2AM he he'd still be there on the opening statement speaking to an empty auditorium
You are too dumb of a racist troll to understand that it's you who is doing damage to the forum/ thread. It's you who C/P pseudo blogs and everything in support of your Eurocentric notions.
It's me who posts peer reviewed sources to debunk your claims and support, as I always do, you fake African American woman, impostor and racist.
Posts: 22244 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
TP's method of argumention is to swamp the issue with mega posts. You start a thread and he starts a new thread within your thread at the same time (he's trying to compete with zarahan) Like if this was a debate at a university one guy would would have a 15 minute opening statement and then Troll would come on and at 2AM he he'd still be there on the opening statement speaking to an empty auditorium
I agree. Zaharan etc., is too much. At least if you're going to use a paper to support your argument paraphrase it and leave a link to the study so others can look at it and see if your interpretation is correct then let an adult debate be had. The spam is just too much and derails threads.
Posts: 46 | From: Los Angeles | Registered: Oct 2010
| IP: Logged |






 UBBFriend: Email this page to someone!
UBBFriend: Email this page to someone!

















![[Big Grin]](biggrin.gif)







 Printer-friendly view of this topic
Printer-friendly view of this topic