Chad Genetic Diversity Reveals an African History Marked by Multiple Holocene Eurasian Migrations
Marc Haber1, , , Massimo Mezzavilla1, 2, Anders Bergström1, Javier Prado-Martinez1, Pille Hallast1, 3, Riyadh Saif-Ali4, Molham Al-Habori4, George Dedoussis5, Eleftheria Zeggini1, Jason Blue-Smith6, 10, R. Spencer Wells7, Yali Xue1, Pierre A. Zalloua8, 9, Chris Tyler-Smith1, , Show more http://dx.doi.org/10.1016/j.ajhg.2016.10.012
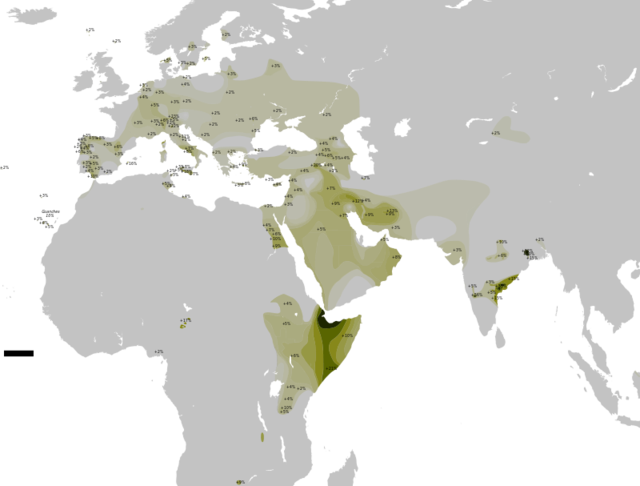
Understanding human genetic diversity in Africa is important for interpreting the evolution of all humans, yet vast regions in Africa, such as Chad, remain genetically poorly investigated. Here, we use genotype data from 480 samples from Chad, the Near East, and southern Europe, as well as whole-genome sequencing from 19 of them, to show that many populations today derive their genomes from ancient African-Eurasian admixtures. We found evidence of early Eurasian backflow to Africa in people speaking the unclassified isolate Laal language in southern Chad and estimate from linkage-disequilibrium decay that this occurred 4,7507,200 years ago. It brought to Africa a Y chromosome lineage [R1b-V88] whose closest relatives are widespread in present-day Eurasia; we estimate from sequence data that the Chad R1b-V88 Y chromosomes coalesced 5,7007,300 years ago. This migration could thus have originated among Near Eastern farmers during the African Humid Period. We also found that the previously documented Eurasian backflow into Africa, which occurred ∼3,000 years ago and was thought to be mostly limited to East Africa, had a more westward impact affecting populations in northern Chad, such as the Toubou, who have 20%30% Eurasian ancestry today. We observed a decline in heterozygosity in admixed Africans and found that the Eurasian admixture can bias inferences on their coalescent history and confound genetic signals from adaptation and archaic introgression.
Results Genetic Structure in Chad Indicates a Complex Admixture History
We performed an initial exploration of our dataset by using principal-component analysis [PCA].19 The first component [PC1] captured the genetic differentiation between Africans and Eurasians [Figure 1B]. Populations such as the Near Easterners and North and East Africans fell between the Europeans and sub-Saharan Africans. The Chadian groups lay near the sub-Saharan Africans: the Sara and Laal speakers clustered tightly with sub-Saharan Africans, such as the Yoruba, whereas the Toubou were somewhat more distant and appeared drawn toward East Africans, such as the Ethiopians. Samples collected from the capital of Chad, NDjamena, appeared in a central position between the Toubou cluster and the Sara and Laal cluster [Figure 1C]. Many individuals from NDjamena have not reported their ethnicity or have reported a mixed ethnic origin. Therefore, recent mixture could be responsible for their position on the PCA.
We further investigated the genetic variation in Chad by estimating changes in the effective populations size [Ne] over time via the MSMC approach.20 Eurasians and Africans diverged around 60,00080,000 ya and subsequently had different patterns of population-size changes: in particular, compared with Africans, the Eurasian population experienced a sharp decrease in size ∼60,000 ya.20 We observed this expected pattern in most populations in our dataset [Figure 2], but a few stood out: [1] Egyptians had a population bottleneck that was much more pronounced than that of other Africans but not as sharp as that of Eurasian populations; and [2] the Toubou and Ethiopians shared a very similar pattern during the bottleneck: they were close to other Africans but had a somewhat sharper decrease in population size [Figure 2]. We would not expect such different fluctuations in population sizes at 60,000 ya in populations who shared a common origin during this period. For example, all Eurasians trace their origin to a population who exited Africa ∼60,000 ya, and this is reflected in indistinguishable Ne patterns during this period, 20 and 33 which we also observed in the CEU, Greeks, and Lebanese [Figure 2], as expected. A shared pattern of Ne in ancient times was also observed in the Sara, Laal speakers, and other Africans, such as the Yoruba. We suggest that the deviation from the expected Ne pattern in the Toubou is related to extensive admixture history with Eurasians, like the Eurasian admixture seen in Ethiopians, and we explore this possibility directly with admixture tests below.
Multiple Eurasian Admixtures in Africa after 6,000 ya
We have previously reported massive gene flow ∼3,000 ya from Eurasians to Ethiopian populations.4 Here, we reassess the presence of Eurasian ancestry in Africa by using f3 statistics25 in the form of f3[X; Eurasian, Yoruba], where a negative value with a Z score < −4 indicates that X is a mixture of Africans and Eurasians. We found, as expected, that most Ethiopians are a mixture of Africans and Eurasians. An exception is the Gumuz population, where f3[Gumuz; Eurasian, Yoruba] is always positive. The Gumuz language belongs to the Nilo-Saharan family, which could have isolated the Gumuz from the Afro-Asiatic-speaking Ethiopians. However, we found that the Toubou in Chad, who also speak a Nilo-Saharan language, are a mixture of Africans and Eurasians, making f3[Toubou; Eurasian, Yoruba] always significantly negative. This suggests that the impact of Eurasian migrations today extends beyond East Africa and the Afro-Asiatic-speaking populations. We did not detect significant [Z score < −4] Eurasian admixture in the Sara [Nilo-Saharan language family] or the Laal speakers [unclassified language] with the use of f3 statistics [lowest Z score for the Sara was >−2.9; for the Laal speakers, Z scores were all positive]. However, this statistic loses sensitivity with small mixture proportions and post-admixture drift, 27 so positive values from the f3 statistics do not necessarily reflect a complete absence of admixture. We thus further tested for admixture by using ALDER and MALDER, which assess admixture-induced linkage disequilibrium [LD] and can detect small mixture proportions from a substantially diverged reference possibly missed by the f3 statistic. ALDER detected admixture in the Toubou, Sara, and Laal speakers [Table S2]. MALDER, which has the potential to determine whether or not the admixture LD in the population is best represented as the result of one or multiple mixtures, showed that two mixture events had occurred in the Toubou [Figure 3A; Table S3]. The first event occurred 2,8503,500 ya [Z score = 11], a time close to the date of mixture in East Africans 2,5002,700 ya [Z score = 26]. The second mixture event occurred much more recently at 170260 ya [Z score = 5]. In southern Chad, we detected mixture events that were more ancient than those in the north. Mixture occurred 3,9004,800 ya [Z score = 10] in the Sara and 4,7507,200 ya [Z score = 5] in the Laal speakers [ Figure 3A]. These time estimates overlap, and we interpret them as signals from the same admixture event, whose time in the distant past was estimated more reliably in the Laal speakers because they carry more Eurasian ancestry [1.25%4.5%] than the Sara [0.3%2%] [see estimates of admixture proportions below], even though the Sara have smaller standard errors because of their larger sample size. In particular, we suggest that the Eurasian mixture event in the Sara and Laal speakers is independent of the mixture event in East Africans and the Toubou for two reasons: [1] admixture LD showed that the events in southern Chad preceded the events in East Africa by 2,0004,500 years, and [2] we found in Chad a Eurasian Y chromosome lineage [Y haplogroup R1b-V88] that had penetrated all Chadian populations examined but was absent or rare from the Ethiopians examined [Table S4; Figure S1]. From whole Y chromosome sequences [Figure S2], we estimate that the Chadian R1b-V88 chromosomes sampled emerged 5,7007,300 ya [Figure 3B], a time comparable to the Laal speaker admixture dates [4,7507,200 ya] estimated from genome-wide LD-decay patterns.
The Sources of Eurasian Backflow into Chad and East Africa Are Correlated
Previous studies have suggested that the Eurasian backflow into East Africa came from a population related to early Neolithic farmers.5 We wanted to know whether the Eurasian ancestry we found in the Toubou, which we attribute to a mixture close in time to the date of mixture in East Africans, can be traced to the same source populations that influenced Ethiopia. We performed the tests f3[Toubou; Yoruba, X] and f3[Amhara; Yoruba, X], where X is a present-day non-sub-Saharan African population in our dataset and is related to one that contributed ancestry to the Toubou and Amhara [Z score < −4] [ Table S5]. We then looked at the correlation of the f3 statistic values between the two tests [Figure 4A]. We found that the Eurasian source populations for the Amhara and Toubou were highly correlated [r = 0.98; 95% CI = 0.980.99; p value < 2.2 × 10−16] and that the most significant result was for present-day Sardinians. Exceptions to this correlation were the North African populations [Tunisians, Mozabite, Algerians, and Saharawi], who appeared to have contributed more ancestry to the Toubou than to the Amhara. We repeated the tests by using published ancient genomes [Table S6] and also found a high correlation of the Eurasian sources for the Amhara and Toubou [r = 0.98; 95% CI = 0.970.99; p value < 2.2 × 10−16]; early Neolithic farmers were the most significant contributors, as reported previously5 [Figure 4B]. When we substituted the Amhara with other Ethiopians [Wolayta and Oromo], we found similar results [data not shown]. In a parallel comparison, we checked whether the sources of the African ancestry in different Near Eastern populations were also correlated. We tested f3[Lebanese; British, X] and f3[Yemeni; British, X] and found a lower correlation of the f3 values [r = 0.62; 95% CI = 0.320.80], suggesting a more complicated history of gene flow from genetically different Africans to different populations in the Near East.
Eurasian Gene Flow Shaped the Genomes of Admixed Africans
Our results from the PCA and MSMC analysis showed a deviation of the admixed populations from the patterns observed in unadmixed [or less admixed] populations in the same geographical region. The MSMC analysis, in particular, showed that admixed Africans had patterns indicative of a decline in heterozygosity [increased bottleneck ∼60,000 ya], somewhat similarly to Eurasians. We tested whole-genome heterozygosity in these populations and found that it decreased in admixed Africans according to their Eurasian ancestry [Figure S4A]. This decrease was not related to recent inbreeding, given that removing segments with long runs of homozygosity did not change the overall pattern. Our simulations suggest that decay in heterozygosity is expected after gene flow from a population with diversity comparable to that of Eurasians [Figures S4B and S4C]. We further investigated heterozygosity in admixed Africans by assessing heterozygosity of the different ancestral segments in the Toubou genome. We found that admixed African-Eurasian segments had more heterozygosity [1.23 hets/kb] than segments of the genome where African-African haplotypes were present [1.19 hets/kb] [Figure S5]. However, the Toubou genome segments with complete Eurasian ancestry [Eurasian-Eurasian] had considerably lower heterozygosity [∼0.96 hets/kb; Figure S5], leading to the genome-wide pattern of decay in heterozygosity observed in Africans with Eurasian ancestry [Figure S4].
We wanted to understand the consequence of admixture on the models that use the density of heterozygous sites to infer the demographic history of populations. We first tested whether the coalescent history estimated by MSMC was affected by a small proportion of mixture, such as the African mixture found in Greeks and Lebanese [ranging from 0% to 5%]. We tested the Greek, Lebanese, CEU, and CHB [Han Chinese in Beijing, China] split times from the Yoruba and found that all populations split from the Yoruba ∼70,00080,000 ya, implying that the low proportions of African admixture in the Greeks and Lebanese did not detectably affect the estimates of relative cross-coalescence rate [Figure S6A]. We next tested the Toubou, who have ∼30% Eurasian ancestry. The Toubou appeared to split from Eurasians ∼30,00040,000 ya, a time more recent than expected considering the African-Eurasian split 60,00080,000 ya20 [Figure S6B]. We tested other Africans in our dataset and found that the Sara, Laal speakers, and Yoruba split from Eurasians, as expected, ∼70,00080,000 ya [Figure S6B]. We then tested directly whether the Eurasian ancestry affected the relative cross-coalescence rate between the Toubou and Eurasians by masking some of the Eurasian ancestry in the Toubou. We used PCAdmix21 to estimate the ancestry along each chromosome and then used the identified Eurasian segments as a negative mask in our analysis. The split times between the Toubou and Lebanese, for example, increased by ∼15,000 years [Figure S6B], shifting the split date toward the expected African-Eurasian split time.
Discussion We have generated an extensive set of genotyping and high-coverage whole-genome sequencing data to study the genetic history of Chad and neighboring populations. We found substantial genetic differences between the ethnic groups inhabiting Chad today and suggest that multiple ancient Eurasian migrations played a major role in shaping the genetic diversity of the region [Figure 3C]. Here, we discuss these migrations and how the mixed ancestry can confound proper interpretation of the evolutionary processes that occurred in their history and therefore needs to be thoroughly accounted for in the study of African genetic diversity.
We detected the earliest Eurasian migrations to Africa in the Laal-speaking people, an isolated language group of fewer than 800 speakers who inhabit southern Chad. We estimate that mixture occurred 4,7507,200 ya, thus after the Neolithic transition in the Near East, a period characterized by exponential growth in human population size. Environmental changes during this period [which possibly triggered the Neolithic transition] also facilitated human migrations. The African Humid Period, for example, was a humid phase across North Africa that peaked 6,0009,000 ya37 and biogeographically connected Africa to Eurasia, facilitating human movement across these regions.38 In Chad, we found a Y chromosome lineage [R1b-V88] that we estimate emerged during the same period 5,7007,300 ya [Figure 3B]. The closest related Y chromosome groups today are widespread in Eurasia and have been previously associated with human expansions to Europe.39 and 40 We estimate that the Eurasian R1b lineages initially diverged 7,3009,400 ya, at the time of the Neolithic expansions. However, we found that the African and Eurasian R1b lineages diverged 17,90023,000 ya, suggesting that genetic structure was already established between the groups who expanded to Europe and Africa. R1b-V88 was previously found in Central and West Africa and was associated with a mid-Holocene migration of Afro-asiatic speakers through the central Sahara into the Lake Chad Basin.8 In the populations we examined, we found R1b in the Toubou and Sara, who speak Nilo-Saharan languages, and also in the Laal people, who speak an unclassified language. This suggests that R1b penetrated Africa independently of the Afro-asiatic language spread or passed to other groups through admixture.
In addition to the early Eurasian migration to Africa ∼6,000 ya, a second migration ∼3,000 ya affected the Toubou population in northern Chad but had no detectable genetic impact on other Chadian populations. This migration appears to be associated with the previously reported Eurasian backflow into East Africa, given that the source populations and dates of mixture are similar. Occurring at the start of the Iron Age, these migrations could have been facilitated by advances in warfare and transportation technology in the Near East. It is uncertain why the impact of this migration in Chad affected only the Toubou. The African ancestral component in the Toubou is best represented by the Laal-speaking population, suggesting that the African-Eurasian mixture probably occurred in Chad. However, ethnolinguistic barriers could have already been established at this time between the Chad groups, preventing a widespread dissemination of the Eurasian ancestry. The Toubou, despite their Islamic faith, do not show the genetic admixture detected in many Near Eastern and North African populations around 1,100 ya,41 suggesting conversion without population mixing at this time. They did, however, receive additional Eurasian ancestry in the past 200 years from a source represented by North African populations such as Tunisians, Mozabite, Algerians, and Sahrawi [Figure 3C]. This recent interaction could have been promoted by the nomadic lifestyle of the present-day Toubou and a shared Muslim religion with North Africans. Unsurprisingly, we also detected a likely mixing of Chad populations in the sample from the capital, which could be even more recent.
Eurasian backflow into Africa thus appears to have been a recurrent event in the history of many Africans, given its considerable impact on their genomes. Although population mixture in general is a process that increases genetic diversity, we observed a decrease in heterozygosity in the admixed Africans. Our simulations showed that these results are expected after mixture at these proportions with the Eurasians who suffered a significant bottleneck at the time of their exodus from Africa ∼60,000 ya. Consequently, we found that mixture can complicate interpretation of the coalescent history inferred from models that use the density of heterozygous sites in their implementations. In addition, we detected in admixed Africans an inflation of positive-selection signals on alleles associated with adult lactose tolerance and pigmentation in Europeans, but we suggest that these alleles have drifted neutrally in Africans after admixture. Furthermore, we detected Neanderthal ancestry in admixed Africans and found it to be proportional to their Eurasian ancestry. Similarly, in admixed Near Easterners, we found a decrease in Neanderthal ancestry proportional to the gene flow they have received from Africans. Although a higher genetic affinity of Neanderthals to Europeans than to Near Easterners was previously interpreted as additional Neanderthal admixture in the history of Europeans,42 we propose that a more parsimonious explanation for these observations is that African-Eurasian mixtures both introduced Neanderthal ancestry to Africa and diluted the Neanderthal ancestry in the Near East.
It is important to note that in this work we inevitably invoke Occams razor to support the simplest model consistent with our data; the history of the populations studied here, including the time and sources of the Eurasian admixture in Africa, could be more complex. aDNA from Chad and neighboring regions remains a challenge given the poor DNA preservation in hot climates, but future successful efforts in aDNA research could provide additional insights and reveal additional complexities not considered by the modern-DNA-based models favored here.43
Our study has shown that human genetic diversity in Africa is still incompletely understood and that ancient admixture adds to its complexity. This work highlights the importance of exploring underrepresented populations, such as those from Chad, in genetic studies to improve our understanding of the demographic processes that shaped genetic variation in Africa and globally.
Acknowledgments We thank all sample donors for making this work possible and the Wellcome Trust Sanger Institute pipelines for generating genotype and sequence data. We also thank Andrea Massaia for comments on analyzing Y chromosome haplogroups from array data and David Soria-Hernanz for comments on the ethnic groups and languages in Chad. M.H., M.M., A.B., J.P.-M., E.Z., Y.X., and C.T.-S. were supported by the Wellcome Trust [098051]. P.H. was supported by Estonian Research Council grant PUT1036. J.B.-S. is a full-time employee of Karius, Inc., and R.S.W. is founder and CEO of Insitome.
Posted by ELIMU (Member # 21677) on :
V88 was most likely brought to Africa by Phoenicians or sidonians. I tend to believe that Chadic speakers came from Sidon in northern Canaan,present day Levant. I also believe that Fula,peul,Fulani,woodabe came from Northern Libya and are most likely mixed with Phoenicians,unlike other tribe of Ancient north Libyans eg Serer,wolof and Libu of present day Senegambia.
Posted by Clyde Winters (Member # 10129) on :
This paper is a joke. It makes numerous speculations without any archaeological evidence to back it up. It claims that R1 in Africa is the result of a back migration, and the Eurasians and Africans diverdge 60-80kya. This dating is contradictory, because the only anatomically modern humans at this time were in Africa, and we know that there is no archaeological evidence of Eurasians migrating into Africa.
The authors "estimate from sequence data that the Chad R1b-V88 Y chromosomes coalesced 5,7007,300 years ago" during a migration from the Levant back into Africa. This was impossible because the Levant at this time was mainly occupied by Sub-Saharan Africans.
Trenton W. Holliday, tested the hypothesis that if modern Africans had dispersed into the Levant from Africa, "tropically adapted hominids" would be represented in the archaeological history of the Levant, especially in relation to the Qafzeh-Skhul hominids. This researcher found that the Qafzeh-Skhul hominids (20,000-10,000),were assigned to the Sub-Saharan population, along with the Natufians samples (4000 BP). Holliday also found African fauna in the area.
If the population was sub-saharan african and the flora and fauna was Sub-Saharan African in the Levant 7,300-4000 BP, the only people who would have made a back migration to Chad and Ethiopia during the proposed back migration of the authors would have been Africans, not Eurasians because their were no Eurasians living in the Levant.
The idea that V88 originated in Eurasia and later deposited in Chad by Eurasians lacked any foundation in the first place. It lacked any foundation because V88 was supposed to be confined mainly to Africa.
This article was another attempt to make it appear that R1 originated in Europe instead of Africa. The dates in this paper are all Eurocentric speculation meant to deny Africans currency in their own development.
Posted by beyoku (Member # 14524) on :
quote:Originally posted by Clyde Winters: This paper is a joke...........................
Trenton W. Holliday, tested the hypothesis that if modern Africans had dispersed into the Levant from Africa, "tropically adapted hominids" would be represented in the archaeological history of the Levant, especially in relation to the Qafzeh-Skhul hominids. This researcher found that the Qafzeh-Skhul hominids (20,000-10,000),were assigned to the Sub-Saharan population, along with the Natufians samples (4000 BP)
Here is Holliday analysis of the Natufian
They dont group with Africans. Not sure what limb proportion data you are looking at.
Posted by the lioness, (Member # 17353) on :
quote:
The Ain Dokhara specimen also falls within the scatter of recent African means. In contrast, none of the Afalou specimens, nor of the El Wad Natufian specimens, falls within the African scatter, and all lie toward the more cold-adapted end of the scatter.
Conclusions
....The Jebel Sahaba sample are distinct in body shape from penecontemporary humans from Afalou-Bou- Rhummel (Algeria) and El Wad Natufians from the southern Levanta result consistent with the results of both Irish (2000, 2005) using dental data and Franciscus (1995, 2003) using nasal data.
--2013 DOI: 10.1002/oa.2315 Population Affinities of the Jebel Sahaba Skeletal Sample: Limb Proportion Evidence T. W. HOLLIDAY*
Posted by the lioness, (Member # 17353) on :
Haplogroup P1 (P-M45), the immediate ancestor of Haplogroup R, likely emerged in South East Asia.[3] The SNP M207, which defines Haplogroup R, is believed to have arisen during the Upper Paleolithic era, about 27,000 years ago.[2][1]
Only one confirmed example of basal R* has been found, in 24,000 year old remains, known as MA-1, found at Mal'ta near Lake Baikal in Siberia.[2] While a living example of R-M207(xM17,M124) was reported in 2012, the sample of 158 ethnic Tajik males from Badakshan, Afghanistan were not tested for the SNP M478; the male concerned may therefore belong to R2*.
Despite the rarity of R*, the relatively rapid expansion geographically and numerically of subclades from R1 in particular, was noted in a 2014 paper by Pille Hallast, Chiara Batini, Daniel Zadik and others: "both R1a and R1b comprise young, star-like expansions" (Karafet 2008).
Posted by DD'eDeN (Member # 21966) on :
the lioness,
Note: Laal is as yet unclassified Boyeldieu (1982b): Its classification remains problematic; while it shows certain common lexical, and no doubt morphological, traits with the Bua languages(Adamawa-13,Niger-Congo family of Joseph H. Greenberg), it differs from them radically in many ways of which some, a priori, are reminiscent of geographically nearby Chadic languages"
Is Bua related to Mbo (oldest Y DNA 300ka)? I would say yes, both refer to mother/birth, related to Mabul(Hebrew)-swell, mene (Egypt) - month, buat (Malay) make-birth. In Javanese, Mbo means mother. - - - The westward migrants may have been pushed by people of the Black Sea oasis due to 7,700 deluge.
Posted by Clyde Winters (Member # 10129) on :
Trenton W. Holliday (2000), tested the hypothesis that if modern Africans had dispersed into the Levant from Africa, "tropically adapted hominids" would be represented in the archaeological history of the Levant, especially in relation to the Qafzeh-Skhul hominids. This researcher found that the Qafzeh-Skhul hominids (20,000-10,000),were assigned to the Sub-Saharan population, along with the Natufians samples (4000 BP). Holliday also found African fauna in the area. Below are a few quotes from the paper by Holliday they show that the population at this time were Negroid in Southwest Asia.
"In this light, some of the more robust assignments (albeit not 95% of the Qafzeh-Skhul hominids to the sub-Saharan African sample (e.g., Qafzeh 8 at 85%, Skhul 4 at 71%) are remarkable indeed" (p. 62).
"The Qafzeh-Skhul hominids have sometimes been refered to as "Proto-CroMagnons" (e.g., Howell 1957; Vandermeersch 1996) because of their presumed similarity to the famous Aurignacian-associated hominids from Western Europe....Specifically [Brace], he notes that "in both the details of its dental and craniological size and from Qafzeh is an unlikely proto-Cro-Magnon, but it makes a fine model for the ancestors of modern sub-Saharan Africans"(p.63).
"taken as a whole, the work of Tchernov seems to support the findings of the current research that the Qafzeh-Skhul hominids have their origins in Africa, while the Neanderthals are from cold to temperate biomes"(p.64).
"The current study demonstrates African-like affinities in the body shape of the Qafzeh-Skhul hominids. This finding is consistent with craniofacial evidence (Brace 1996) and with zooarchaeological data indicating the presence of African fauna at Qafzeh (Rabinovich and Tchernov 1995; Tchernov 1988, 1992)" (p.64).\
Holiday, T. (2000). Evolution at the Crossroads: Modern Human Emergence in Western Asia, American Anthropologist,102(1) . Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by ELIMU: V88 was most likely brought to Africa by Phoenicians or sidonians. I tend to believe that Chadic speakers came from Sidon in northern Canaan,present day Levant. I also believe that Fula,peul,Fulani,woodabe came from Northern Libya and are most likely mixed with Phoenicians,unlike other tribe of Ancient north Libyans eg Serer,wolof and Libu of present day Senegambia.
lol
So, on what do you base the Phoenicians were of Hg R*?
quote: How does the present MSY tree compare with the backbone of the recently published reference MSY phylogeny?13 The phylogenetic relationships we observed among chromosomes belonging to haplogroups B, C, and R are reminiscent of those reported in the tree by Karafet et al.13
--Fulvio Cruciani et al A Revised Root for the Human Y Chromosomal Phylogenetic Tree: The Origin of Patrilineal Diversity in Africa (2011)
quote: New work [2022] has addressed the Neolithic transition in Europe by focusing on the main western European Y chromosome haplogroup R1b1b2-M269 (hereafter referred to as R-M269). This lineage had hitherto received little recent attention in this context, although previous work suggested that the broader R-M173 clade (excluding the R1a-M17 sub-lineage) and Haplogroup 1 (derived at single nucleotide polymorphism, or SNP, 92r7) are likely to have spread into Europe during the Palaeolithic [17,18,23], and therefore unlikely to have been carried into Europe with the migrating farmers. Balaresque et al.
--George B. J. Busby
The peopling of Europe and the cautionary tale of Y chromosome lineage R-M269
Published 25 January 2012.DOI: 10.1098/rspb.2011.1044
quote:
This branching pattern, along with the geographical distribution of the major clades A, B, and CT, has been interpreted as supporting an African origin for anatomically modern humans,10 with Khoisan from south Africa and Ethiopians from east Africa sharing the deepest lineages of the phylogeny.15 and 16
[...]
The deepest branching separates A1b from a monophyletic clade whose members (A1a, A2, A3, B, C, and R) all share seven mutually reinforcing derived mutations (five transitions and two transversions, all at non-CpG sites).
[...]
How does the present MSY tree compare with the backbone of the recently published reference MSY phylogeny?13 The phylogenetic relationships we observed among chromosomes belonging to haplogroups B, C, and R are reminiscent of those reported in the tree by Karafet et al.13 These chromosomes belong to a clade (haplogroup BT) in which chromosomes C and R share a common ancestor (Figure 2).
quote: The Y chromosome Alu polymorphism (YAP, also called M1) defines the deep-rooted haplogroup D/E of the global Y-chromosome phylogeny [1]. This D/E haplogroup is further branched into three sub-haplogroups DE*, D and E (Figure 1). The distribution of the D/E haplogroup is highly regional, and the three subgroups are geographically restricted to certain areas, therefore informative in tracing human prehistory (Table 1). The sub-haplogroup DE*, presumably the most ancient lineage of the D/E haplogroup was only found in Africans from Nigeria [2], supporting the "Out of Africa" hypothesis about modern human origin. The sub-haplogroup E (E-M40), defined by M40/SRY4064 and M96, was also suggested originated in Africa [3-6], and later dispersed to Middle East and Europe about 20,000 years ago [3,4]. Interestingly, the sub-haplogroup D defined by M174 (D-M174) is East Asian specific with abundant appearance in Tibetan and Japanese (3040%), but rare in most of other East Asian populations and populations from regions bordering East Asia (Central Asia, North Asia and Middle East) (usually less than 5%) [5-7]. Under D-M174, Japanese belongs to a separate sub-lineage defined by several mutations (e.g. M55, M57 and M64 etc.), which is different from those in Tibetans implicating relatively deep divergence between them [1]. The fragmented distribution of D-M174 in East Asia seems not consistent with the pattern of other East Asian specific lineages, i.e. O3-M122, O1-M119 and O2-M95 under haplogroup O [8,9].
quote: Further refinement awaits the finding of new markers especially within paragroup E3a*-M2. The microsatellite profile of the DE* individual is one mutational step away from the allelic state described for Nigerians (DYS390*21, DYS388 not tested; [37], therefore suggesting a common ancestry but not elucidating the phylogenetics.
Haplogroup DE* in Guinea-Bissau:
Y-chromosomal diversity in the population of Guinea-Bissau: a multiethnic perspective
quote: There has been considerable debate on the geographic origin of the human Y chromosome Alu polymor- phism (YAP). Here we report a new, very rare deep-rooting haplogroup within the YAP clade, together with data on other deep-rooting YAP clades. The new haplogroup, found so far in only five Nigerians, is the least-derived YAP haplogroup according to currently known binary markers. However, because the interior branching order of the Y chromosome genealogical tree remains unknown, it is impossible to impute the origin of the YAP clade with certainty. We discuss the problems presented by rare deep-rooting lineages for Y chromosome phylogeography.
Haplogroup DE* in Nigerians:
Rare Deep-Rooting Y Chromosome Lineages in Humans: Lessons for Phylogeography
quote: Figure 3: Evidence of gene flow from a population related to MA-1 and western Eurasians into Native American ancestors.
From Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans
Maanasa Raghavan, Pontus Skoglund, Kelly E. Graf, Mait Metspalu, Anders Albrechtsen, Ida Moltke, Simon Rasmussen, Thomas W. Stafford Jr, Ludovic Orlando, Ene Metspalu, Monika Karmin, Kristiina Tambets, Siiri Rootsi, Reedik Mägi, Paula F. Campos, Elena Balanovska, Oleg Balanovsky, Elza Khusnutdinova, Sergey Litvinov, Ludmila P. Osipova, Sardana A. Fedorova, Mikhail I. Voevoda, Michael DeGiorgio, Thomas Sicheritz-Ponten, Søren Brunak et al.
Nature (2013) doi:10.1038/nature12736
Allele frequency-based D-statistic tests20 of the forms. a, D(Yoruba, MA-1; Han, X), where X represents modern-day populations from North and South America. The D-statistic is significantly positive for all the tests, providing evidence for gene flow between Native American ancestors and the MA-1 population lineage; however, it is not informative with respect to the direction of gene flow. b, D(Yoruba, X; Han, Karitiana), where X represents non-African populations. Since all of the 17 tested western Eurasian populations are closer to Karitiana than to Han Chinese, the most parsimonious explanation is that Native Americans have western Eurasian-related ancestry. c, D(Sardinian, X; Papuan, Han), where X represents non-African populations. MA-1 is not significantly closer to Han Chinese than to Papuans, which is compatible with MA-1 having no Native American-related admixture in its ancestry. Thick and thin error bars correspond to 1 and 3 standard errors of the D-statistic, respectively.
Admixture graphs were constructed relating MA-1 to modern groups using MixMapper v1.01, a recently developed approach that utilizes f2 distances between populations to fit admixture graphs of population history. MixMapper is based on an approach where a scaffold tree is constructed from populations that are assumed to be unadmixed, upon which other populations can then be fitted as either simple branches off the scaffold, two-way admixtures between populations in the scaffold tree, or alternatively three-way admixtures between an unadmixed population, and a two-way admixed population.
In order to place the MA-1 genome in the context of modern day variation, a scaffold tree was constructed using 4 African genomes (San, Yoruba, Mandenka, Dinka), Sardinian and Han3. The Mbuti genome was excluded for the main analyses since the Mbuti are known to have substantial amounts of recent Bantu-speaker-related ancestry, and are difficult to fit using admixture graphs1,2, but similar results were obtained by us when the Mbuti was also included (see below). Sardinian and Han were chosen as representatives of the western and eastern extremes of modern-day Eurasian genetic differentiation. All transitions were excluded, and standard errors of the f-statistics were estimated using 500 bootstrap replicates over 50 blocks of the autosomal genome
When adding the Karitiana genome to this scaffold tree, 422 of 500 bootstrap pseudoreplicates were found to fit Karitiana as a mix between the Sardinian and Han lineages, with 73.7% (95% CI: 61.8-85.7%) of its ancestry being derived from the Han lineage and the remainder 26.3% (14.3-38.2%) from the Sardinian lineage (Figure SI 18; Table SI 12). When the Mbuti was included, the bootstrap support was 432 of 500 and the Sardinian-related ancestry in MA-1 was 13.4-35.4%. Fitting MA-1 to the scaffold with Sardinian, the best fit (496 of 500 bootstraps) suggested that MA- 1 is a mixture of the Sardinian and Han lineages, with 22.1-75.0% of its ancestry being derived from the Han lineage (Figure SI 18; Table SI 12).
Subsequently, Karitiana was fitted as a three-way mixture between MA-1 (modeled as a mixture as above, see SI 14) and Han, since it is likely that populations related to MA-1 mediated the mixture event between western Eurasians and Native Americans. Here, 496 bootstrap replicates supported Karitiana as having 26.1% (7.7-44.4%) ancestry from the MA-1 lineage and the remainder from Han, consistent with the previous analysis.
A previous study1 used MixMapper and data ascertained in a San individual, and subsequently genotyped in a large number of human populations, to fit Sardinians as a mixture of a western Eurasian lineage and Karitiana, which would suggest that it is inappropriate to use Sardinians in the scaffold tree. If the Sardinian is replaced with Karitiana in the scaffold tree, the highest number of bootstrap pseudoreplicates for any one combination (231 of 500) fitted Sardinians as a mix between Dinka and Karitiana (with 27.7-33.4% ancestry from the Dinka lineage). Lipson et al. (2013) 1 chose the set of populations to be included in the scaffold based on observed f3 tests and additivity of f2-distances in the scaffold tree. However, as discussed later on in SI 14, the reason for Native Americans not displaying negative f3-statstics when tested for having dual ancestry from eastern and western Eurasians could be the extensive amount of genetic drift that has occurred since their divergence. We found that the scaffold tree with Sardinian was more additive than the scaffold tree with Karitiana, suggesting that Karitiana are better fitted as admixed than Sardinians are (Figure SI 19). This was also consistent when Han was replaced by Dai in the scaffold tree and when both East Asians were included (Figure SI 19). It was also consistent when Mbuti was included in the scaffold tree for either (or both) East Asian genomes.
Sardinians also do not display any significant f3-tests, or evidence of admixture linkage disequilibrium with Native Americans4,5. However, we note that if Sardinians themselves have admixture from a Native American source as suggested by Lipson et al. (2013)1, or some other type of complex ancestry (they have evidence of 0.6±0.2% African ancestry5,6 but that is likely negligible for our estimates), the estimates of western Eurasian ancestry in Karitiana are likely to be affected.
Current admixture graph models are not suitable for dealing with possible bidirectional gene flow, and so a future line of study would be to simultaneously infer gene flow between genomic samples from Native American and western Eurasian populations.
Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans
Nature 505, 8791 (02 January 2014) doi:10.1038/nature12736
Received 14 July 2013 Accepted 04 October 2013 Published online 20 November 2013
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by the lioness,:
quote:
The Ain Dokhara specimen also falls within the scatter of recent African means. In contrast, none of the Afalou specimens, nor of the El Wad Natufian specimens, falls within the African scatter, and all lie toward the more cold-adapted end of the scatter.
Conclusions
....The Jebel Sahaba sample are distinct in body shape from penecontemporary humans from Afalou-Bou- Rhummel (Algeria) and El Wad Natufians from the southern Levanta result consistent with the results of both Irish (2000, 2005) using dental data and Franciscus (1995, 2003) using nasal data.
--2013 DOI: 10.1002/oa.2315 Population Affinities of the Jebel Sahaba Skeletal Sample: Limb Proportion Evidence T. W. HOLLIDAY*
Interesting is:
quote: These results corroborate those of both Irish and Franciscus, who, using dental, oral and nasal morphology,[...]
The body shape of the terminal Pleistocene Jebel Sahaba population is tropical-adapted, with elongated limbs, especially in the distal segments, and is most similar to living sub-Saharan Africans and less similar to late Pleistocene and Holocene North Africans (including Egyptians and Nubians).[...]
Of which sub-Saharan Africans do that speak?
Posted by beyoku (Member # 14524) on :
@ Clyde.....don't be stupid. You said African in reference to tropical adaptation. negroid = cranial facial. Limbs = Tropical adaptation. Where did you reference tropical adaptation for Natufian?
Posted by Swenet (Member # 17303) on :
quote:This researcher found that the Qafzeh-Skhul hominids (20,000-10,000),were assigned to the Sub-Saharan population, along with the Natufians samples (4000 BP).
Clyde will always get a special respect from me because he and Rashidi (i.e. their writings) literally got me started on this journey. So I rarely go hard on him.
But are we all going act like we didn't notice that those dates are the most funny Clyde blunders we've read in a long time?
Posted by Oshun (Member # 19740) on :
@Ish Gebor, can you explain in very lay people English how the quote responds to lioness, thanks! Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by Oshun: @Ish Gebor, can you explain in very lay people English how the quote responds to lioness, thanks!
Why they had to use 4 African genomes (San, Yoruba, Mandenka, Dinka)" to place the MA-1 genome in the context of modern day variation?
"In order to place the MA-1 genome in the context of modern day variation, a scaffold tree was constructed using 4 African genomes (San, Yoruba, Mandenka, Dinka)"
Posted by Doug M (Member # 7650) on :
Key point from the study:
quote: Our study has shown that human genetic diversity in Africa is still incompletely understood and that ancient admixture adds to its complexity. This work highlights the importance of exploring underrepresented populations, such as those from Chad, in genetic studies to improve our understanding of the demographic processes that shaped genetic variation in Africa and globally.
Meaning 1) Chad as one of the nations straddling the Sahara and Sahel should be part of any genetic study of "North African" populations, but for some reason (sampling bias), it is not. 2) If North Africans are supposedly "admixed" with ancient "Eurasians" then what about populations on the border of the Sahara? Are they too mixed? and 3) Ultimately this all boils down to trying to decide whether the Sahara was an ancient conduit for Africans migrating INTO Europe or was it a gateway for back migration FROM Europe. And my opinion is that Europeans are still too racist to admit that their ancestors in Southern Europe in the past had significant African ancestry. So it is the same old game being played with biased and sloppy sample sets and skewed results to support an a-priori assumption. The fundamental gist of this is to rewrite human history into some history of European migrations, as opposed to the fact that human history is the history of African migrations around the planet.
This stuff sounds no different than the ancient brown race European origin of "brown" Africans crap.
Posted by the lioness, (Member # 17353) on :
Posted by Djehuti (Member # 6698) on :
Is it just me, or do the graphics of the paper seem to suggest if not strongly imply that the very origins of Afrasian langauge phylum is Eurasian in origin.
By the way, something the paper fails to mention is why it is not only R1b-V88 but also R1* occurr at high frequency in Cameroonians who don't speak Afrasian at all but Niger-Congo, even though Lioness presented a paper here claiming Cameroonians showed no admixture. So what does this mean??
Posted by the lioness, (Member # 17353) on :
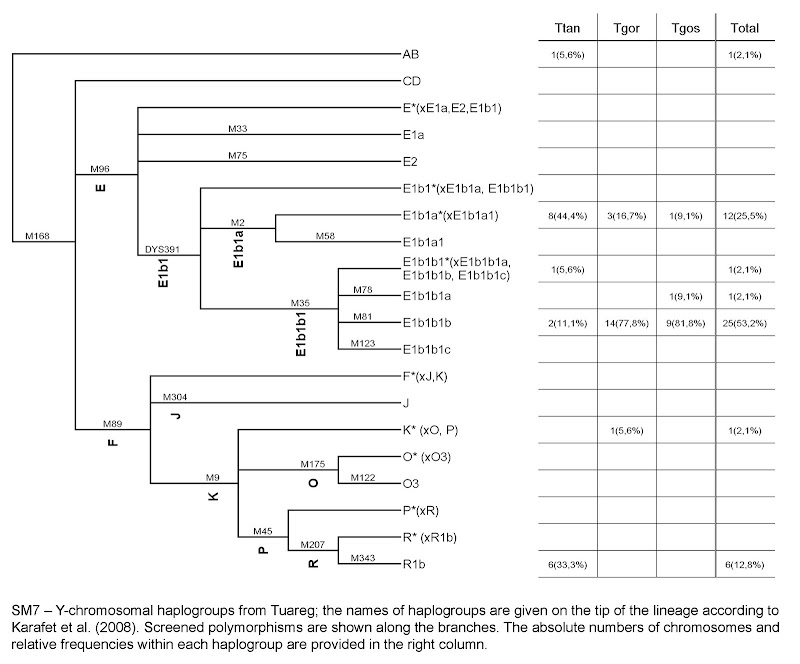
Figure 3. Timing of the Eurasian Admixture in Africa (A) Crosses represent significant admixture events in the history of the Toubou, Amhara, Sara, and Laal speakers. Time of admixture is estimated from LD by ALDER with all pairs of African-Eurasian populations in our dataset as references. MALDER extends ALDER infer- ence by detecting multiple mixture events, such as in the case of the Toubou population (shown here in green lozenges). (B) A maximum-likelihood tree shows the males belonging to haplogroup R1b in the 1000 Genomes Project and the R1b males in our dataset. The number of samples is shown on each branch tip. We estimate that the Chadian R1b emerged 5,7007,300 ya, whereas most European R1b haplogroups emerged 7,3009,400 ya. The African and Eurasian lineages coalesced 17,90023,000 ya. (C) Putative sources and times of admixture of the Eurasian ancestry in Chad and East Africa.
R1b-V88 was previously found in Central and West Africa and was associated with a mid-Holocene migration of Afro-asiatic speakers through the central Sahara into the Lake Chad Basin.8 In the populations we examined, we found R1b in the Toubou and Sara, who speak Nilo-Saharan languages, and also in the Laal people, who speak an unclassified language. This suggests that R1b penetrated Africa independently of the Afro-asiatic language spread or passed to other groups through admixture.
Posted by Doug M (Member # 7650) on :
Here is the key question that people need to be asking and I know this has been discussed here before but it needs to be reiterated. How come African genes outside Africa are quickly labeled as something other than African because of splits, yet so-called Eurasian genes in Africa are Eurasian even thousands of years later and after multiple splits?
See the pattern here? Multiple papers in a short time period talking of ancient "Eurasian" lineages in Africa, yet Africa is the BASIS of all humans on the planet and there are HARDLY ANY papers on African lineages in Europe, the Levant, Asia and elsewhere. Sounds like a double standard to me.
The following wiki page shows how arbitrary and contradictory the logic goes regarding labeling African genetics versus Eurasian genetics. By the logic they use to label "Eurasian" mixture in Africa, ALL HUMANS outside of Africa and their lineages should be labeled as African for the first 10,000 years of human existence outside of Africa. But note that they don't do this.... Wonder why? Keep in mind that Neanderthal admixture is one scam they use to justify this nonsense.....
Before jumping to critical conclusions about this paper, we should Keep in mind how Soccams razor works or more accurately how 'damage control' works.... On another note I find it incredible how so much supposed reentry is being incrementally discovered. I feel like we're getting worked into a circle. It's only a matter of time before its proposed that in every direction of the continent africans have been mixing with "Eurasians."
Posted by Oshun (Member # 19740) on :
quote:Originally posted by Ish Gebor:
quote:Originally posted by Oshun: @Ish Gebor, can you explain in very lay people English how the quote responds to lioness, thanks!
Why they had to use 4 African genomes (San, Yoruba, Mandenka, Dinka)" to place the MA-1 genome in the context of modern day variation?
"In order to place the MA-1 genome in the context of modern day variation, a scaffold tree was constructed using 4 African genomes (San, Yoruba, Mandenka, Dinka)"
I'm sorry I'm not very familiar with genetics. What is the MA-1 genome and what would the signifficance of the 4 African genomes be??? Posted by the lioness, (Member # 17353) on :
quote:Originally posted by Oshun:
quote:Originally posted by Ish Gebor:
quote:Originally posted by Oshun: @Ish Gebor, can you explain in very lay people English how the quote responds to lioness, thanks!
Why they had to use 4 African genomes (San, Yoruba, Mandenka, Dinka)" to place the MA-1 genome in the context of modern day variation?
"In order to place the MA-1 genome in the context of modern day variation, a scaffold tree was constructed using 4 African genomes (San, Yoruba, Mandenka, Dinka)"
I'm sorry I'm not very familiar with genetics. What is the MA-1 genome and what would the signifficance of the 4 African genomes be???
the Mal'ta people belonged to an extinct population closely related to a population who contributed substantially to the genetic ancestry of Siberians, Native Americans and Bronze Age Yamnaya people. The Yamna or Yamnaya culture, also called Pit Grave Culture and Ochre Grave Culture, was a late Copper Age/early Bronze Age culture of the Southern Russian Pontic steppe, dating to 3,500 2,300 BCE.and believed to have contributed ancestry to modern Europeans
The MA-1 genome is the genome from 24,000-year-old individual (MA-1), from Mal'ta in south-central SiberiaThe MA-1 mitochondrial genome belongs to haplogroup U,
The paternal DNA of the boy was Haplogroup R* the oldest human remains yet found carrying this basal form of haplgroup R (M207) which later split into R1 and R2
Keep in mind that when Ish Gebor doesn't like an article he posts with a bunch of quotes of other articles which are somewhat related to the topic but not necessarily rebutting the topic. You asked him about it and his reply was to ask you another question. That's his usual routine. He can't explain how the articles he posts are a specific reply to the topic. He hopes that the lay people will see all these quotes and think " well look at all the quotes put up he must have disproven something" but if you call him out he can't explain it. This has been going on for years. Hopefully other people will continue actually quote the thread topic article and then comment on it.
Posted by Oshun (Member # 19740) on :
I have a question: What's the distribution of V88 in (modern) Southern Egypt and Sudan?
Posted by Elmaestro (Member # 22566) on :
^Relatively absent, I know that the Sahelian groups such as (Peuls & Hausa) in Sudan carry it moderately, and most other instances can be attributed to contemporary admixture... you can find V88 carriers at moderate frequency in the Siwa residents & Berber people though, but I don't believe that's what you're looking for.
Posted by Oshun (Member # 19740) on :
If V88 carriers settled in Egypt and changed the populations of Egypt/Sudan genetically, why don't we see more of them (unless they didn't)? All the Semetic and European flow that's affected modern Egypt and it's neither in the modern or AE DNA??? It sounds like V88 carriers migrated through Egypt, but didn't mingle w/ the people, nor set up shop there. Sudan's seen a bit more mingling, but it doesn't seem like anything especially major. It'd be nice to have a coalescence date for Sudanese with V88.
Posted by the lioness, (Member # 17353) on :
quote:Originally posted by Oshun: If V88 carriers settled in Egypt and changed the populations of Egypt/Sudan genetically, why don't we see more of them (unless they didn't)? All the Semetic and European flow that's affected modern Egypt and it's neither in the modern or AE DNA??? It sounds like V88 carriers migrated through Egypt, but didn't mingle w/ the people, nor set up shop there. Sudan's seen a bit more mingling, but it doesn't seem like anything especially major. It'd be nice to have a coalescence date for Sudanese with V88.
Like its northern counterpart (R1b-M269), R1b-V88 is associated with the domestication of cattle in northern Mesopotamia. Both branches of R1b probably split soon after cattle were domesticated, approximately 10,500 years ago (8,500 BCE). R1b-V88 migrated south towards the Levant and Egypt. The migration of R1b people can be followed archeologically through the presence of domesticated cattle, which appear in central Syria around 8,000-7,500 BCE (late Mureybet period), then in the Southern Levant and Egypt around 7,000-6,500 BCE (e.g. at Nabta Playa and Bir Kiseiba). Cattle herders subsequently spread across most of northern and eastern Africa. The Sahara desert would have been more humid during the Neolithic Subpluvial period (c. 7250-3250 BCE), and would have been a vast savannah full of grass, an ideal environment for cattle herding.
Evidence of cow herding during the Neolithic has shown up at Uan Muhuggiag in central Libya around 5500 BCE, at the Capeletti Cave in northern Algeria around 4500 BCE. But the most compelling evidence that R1b people related to modern Europeans once roamed the Sahara is to be found at Tassili n'Ajjer in southern Algeria, a site famous pyroglyphs (rock art) dating from the Neolithic era. Some painting dating from around 3000 BCE depict fair-skinned and blond or auburn haired women riding on cows.
After reaching the Maghreb, R1b-V88 cattle herders could have crossed the Strait of Gibraltar to Iberia, probably accompanied by G2 farmers, J1 and T1a goat herders and native Maghreban E-M81 lineages. These Maghreban Neolithic farmers/herders could have been the ones who established the Almagra Pottery culture in Andalusia in the 6th millennium BCE.
Nowadays small percentages (1 to 4%) of R1b-V88 are found in the Levant, among the Lebanese, the Druze, and the Jews, and almost in every country in Africa north of the equator. Higher frequency in Egypt (5%), among Berbers from the Egypt-Libya border (23%), among the Sudanese Copts (15%), the Hausa people of Sudan (40%), the the Fulani people of the Sahel (54% in Niger and Cameroon), and Chadic tribes of northern Nigeria and northern Cameroon (especially among the Kirdi), where it is observed at a frequency ranging from 30% to 95% of men. According to Cruciani et al. (2010) R1b-V88 would have crossed the Sahara between 9,200 and 5,600 years ago, and is most probably associated with the diffusion of Chadic languages, a branch of the Afroasiatic languages. V88 would have migrated from Egypt to Sudan, then expanded along the Sahel until northern Cameroon and Nigeria. However, R1b-V88 is not only present among Chadic speakers, but also among Senegambian speakers (Fula-Hausa) and Semitic speakers (Berbers, Arabs).
R1b-V88 is found among the native populations of Rwanda, South Africa, Namibia, Angola, Congo, Gabon, Equatorial Guinea, Ivory Coast, Guinea-Bissau. The wide distribution of V88 in all parts of Africa, its incidence among herding tribes, and the coalescence age of the haplogroup all support a Neolithic dispersal. In any case, a later migration out of Egypt would be improbable since it would have brought haplogroups that came to Egypt during the Bronze Age, such as J1, J2, R1a or R1b-L23.
The maternal lineages associated with the spread of R1b-V88 in Africa are mtDNA haplogroups J1b, U5 and V, and perhaps also U3 and some H subclades (=> see Retracing the mtDNA haplogroups of the original R1b people
Posted by Swenet (Member # 17303) on :
@Oshun
Sound analysis.
It should be noted, though, that R-V88, being a Y chromosome, is inherited as a single independent unit (meaning, independent from parts of the genome where most of our genetic heritage resides). Because of this, haplogroups don't have to have the affinity baggage associated with closely related haplogroups. So, even though R-V88 is derived from R, a Eurasian haplogroup, we don't know what the R-V88 people flowing into Africa were like, genetically.
In exactly the same way, R-V88 is today associated with Sub-Saharan African versions of sicklemia in some areas:
^So, even IF they find R-V88 in Nile Valley ancient DNA, it doesn't mean it belongs to someone who can be meaningfully called 'Eurasian' or 'non-African'.
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by Djehuti:
Is it just me, or do the graphics of the paper seem to suggest if not strongly imply that the very origins of Afrasian langauge phylum is Eurasian in origin.
By the way, something the paper fails to mention is why it is not only R1b-V88 but also R1* occurr at high frequency in Cameroonians who don't speak Afrasian at all but Niger-Congo, even though Lioness presented a paper here claiming Cameroonians showed no admixture. So what does this mean??
Yes, they cling on to the Alexander Militarev theory.
They want to go back and make it "great again", back to the old reasoning, the way things were.
quote:Originally posted by Doug M: Here is the key question that people need to be asking and I know this has been discussed here before but it needs to be reiterated. How come African genes outside Africa are quickly labeled as something other than African because of splits, yet so-called Eurasian genes in Africa are Eurasian even thousands of years later and after multiple splits?
See the pattern here? Multiple papers in a short time period talking of ancient "Eurasian" lineages in Africa, yet Africa is the BASIS of all humans on the planet and there are HARDLY ANY papers on African lineages in Europe, the Levant, Asia and elsewhere. Sounds like a double standard to me.
The following wiki page shows how arbitrary and contradictory the logic goes regarding labeling African genetics versus Eurasian genetics.
By the logic they use to label "Eurasian" mixture in Africa, ALL HUMANS outside of Africa and their lineages should be labeled as African for the first 10,000 years of human existence outside of Africa. But note that they don't do this.... Wonder why? Keep in mind that Neanderthal admixture is one scam they use to justify this nonsense.....
quote:By the logic they use to label "Eurasian" mixture in Africa, ALL HUMANS outside of Africa and their lineages should be labeled as African for the first 10,000 years of human existence outside of Africa.
Jibril Hirbo, Sara Tishkoff et al.
The Episode of Genetic Drift Defining the Migration of Humans out of Africa Is Derived from a Large East African Population Size
PLoS One. 2014; 9(5): e97674. Published online 2014 May 20. doi: 10.1371/journal.pone.0097674
quote: Colored dots indicate genetic diversity. Each new group outside of Africa represents a sampling of the genetic diversity present in its founder population. The ancestral population in Africa was sufficiently large to build up and retain substantial genetic diversity.
--Brenna M. Henna, L. L. Cavalli-Sforzaa,1, and Marcus W. Feldmanb,2 Edited by C. Owen Lovejoy, Kent State University, Kent, OH, and approved September 25, 2012 (received for review July 19, 2012)
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by Elmaestro: Before jumping to critical conclusions about this paper, we should Keep in mind how Soccams razor works or more accurately how 'damage control' works.... On another note I find it incredible how so much supposed reentry is being incrementally discovered. I feel like we're getting worked into a circle. It's only a matter of time before its proposed that in every direction of the continent africans have been mixing with "Eurasians."
That is exactly the purpose, of Occam's razor. At least if there is a trace of civilization useful to them to claim. This has been a ongoing battle of white supremacy. On has to understand that europeans basically hold monopoly on "(re-)writing history" in their favor.
Ancient DNA and the rewriting of human history: be sparing with Occams razor
Marc Haber, Massimo Mezzavilla, Yali Xue and Chris Tyler-SmithEmail author
quote:Why are other populations of humans so much less genetically varied than Africans? The answer, Henn explains, lies in our ancestors history; *the groups of people that migrated out of Africa and spread throughout other continents were smaller subsets of that original, genetically diverse population.
Brenna Henn: "AND WITHIN EACH OF THESE GROUPS THERE IS AN AMAZING AMOUNT OF DIVERSITY, [...] THE DIVERSITY IS INDIGNIOUS TO AFRICAN POPULATIONS*:
Tracing Family Trees, And Human History, With Genetics
quote:Originally posted by Oshun: @Ish Gebor, can you explain in very lay people English how the quote responds to lioness, thanks!
Why they had to use 4 African genomes (San, Yoruba, Mandenka, Dinka)" to place the MA-1 genome in the context of modern day variation?
"In order to place the MA-1 genome in the context of modern day variation, a scaffold tree was constructed using 4 African genomes (San, Yoruba, Mandenka, Dinka)"
I'm sorry I'm not very familiar with genetics. What is the MA-1 genome and what would the signifficance of the 4 African genomes be???
In order to get to the root of R they had to reconstructed genomes from African ethnic groups. That is what the "supposed random" quotes say. They themselves admitted they couldn't have reconstructed this without the genomes of these African ethnic groups.
Hg R was already in Africa, long before any dispersal. And it was taken outside of Africa by dispersal. For them to admit to this, would crush their entire fallacy.
You have to understand that these was a time the entire Hg E was claimed to be from Eurasia as well. And some still cling on to this.
See, lioness advocates back-migrations from wherever into Africa, whenever as much as possible. With Africans not being able to have civilization or cultivation on their own, only swinging from trees until Eurasians came along to intervene. The moment Africans have dispersal going into Europe, lioness is ready to fight this with tooth and nail. And a major problem with these supped studies is that they make a clear cut distinction on Africa and Eurasian borders. Which is obviously pure politics embedded in old colonial ways.
quote:Originally posted by Oshun: If V88 carriers settled in Egypt and changed the populations of Egypt/Sudan genetically, why don't we see more of them (unless they didn't)? All the Semetic and European flow that's affected modern Egypt and it's neither in the modern or AE DNA??? It sounds like V88 carriers migrated through Egypt, but didn't mingle w/ the people, nor set up shop there. Sudan's seen a bit more mingling, but it doesn't seem like anything especially major. It'd be nice to have a coalescence date for Sudanese with V88.
Like its northern counterpart (R1b-M269), R1b-V88 is associated with the domestication of cattle in northern Mesopotamia. Both branches of R1b probably split soon after cattle were domesticated, approximately 10,500 years ago (8,500 BCE). [...]
LOL How "subjective" (sarc). The part they forgot to write about:
quote:
The site has been directly dated to 9650)9950 calBP (11), showing intense occupation over two to three centuries. The economy of the population has been shown to be that of pastoralists, focusing on goats (11). Archaeobotanical evidence is limited (16) but the evidence present is for two)row barley, probably wild, and no evidence for wheat, rye or other domesticates. In other words the overall economy is divergent from the classic agricultural mode of cereal agriculture found in the Levant, Anatolia and Northern Mesopotamian basin.
[...]
We compared GD13a with a number of other ancient genomes and modern populations (6, 1729), using principal component analysis (PCA) (30), ADMIXTURE (31) and outgroup f3 statistics (32) (Fig. 1). GD13a did not cluster with any other early Neolithic individual from Eurasia in any of the analyses. ADMIXTURE and outgroup f3 identified Caucasus Hunter)Gatherers of Western Georgia, just north of the Zagros mountains, as the group genetically most similar to GD13a (Fig. 1B&C), whilst PCA also revealed some affinity with modern Central South Asian populations such as Balochi, Makrani and Brahui (Fig. 1A and Fig. S4). Also genetically close to GD13a were ancient samples from Steppe populations (Yamanya & Afanasievo) that were part of one or more Bronze age migrations into Europe, as well as early Bronze age cultures in that continent (Corded Ware) (17, 23), in line with previous relationships observed for the Caucasus Hunter)Gatherers (26).
[...]
Figure Legends:
Fig. 1. GD13a appears to be related to Caucasus Hunter Gatherers and to modern South Asian populations.
A) PCA loaded on modern populations (represented by open symbols). Ancient individuals (solid symbols) are projected onto these axes.
B) Outgroup f3(X, GD13a; Dinka), where Caucasus Hunter Gatherers (Kotias and Satsurblia) share the most drift with GD13a. Ancient samples have filled circles whereas modern populations are represented by empty symbols.
C) ADMIXTURE using K=17, where GD13a appears very similar to Caucasus Hunter Gatherers, and to a lesser extent to modern south Asian populations.
quote:Originally posted by Oshun: If V88 carriers settled in Egypt and changed the populations of Egypt/Sudan genetically, why don't we see more of them (unless they didn't)? All the Semetic and European flow that's affected modern Egypt and it's neither in the modern or AE DNA??? It sounds like V88 carriers migrated through Egypt, but didn't mingle w/ the people, nor set up shop there. Sudan's seen a bit more mingling, but it doesn't seem like anything especially major. It'd be nice to have a coalescence date for Sudanese with V88.
The migration of R1b people can be followed archeologically through the presence of domesticated cattle, which appear in central Syria around 8,000-7,500 BCE (late Mureybet period), then in the Southern Levant and Egypt around 7,000-6,500 BCE (e.g. at Nabta Playa and Bir Kiseiba). Cattle herders subsequently spread across most of northern and eastern Africa. The Sahara desert would have been more humid during the Neolithic Subpluvial period (c. 7250-3250 BCE), and would have been a vast savannah full of grass, an ideal environment for cattle herding.
The part they forgot to write about:
quote: Among other groups, the Negroes and Baluch mulattoes of Baluchistan, which now forms part of West Pakistan, are of great interest to students of race and ethnic relations. Negroes in West Pakistan are called Makranis.
[...]
Professor S. K. Chatterji, the Indian linguist, discussing the basic unity underlying the diversity of culture in India, also supports this view. According to him, "the first people to arrive in India were a Negrito or Negroid race from Africa, coming at a very early period by way of Arabia and the coastline of Iran. They spread over western and southern India, and even passed on to the northeastern part of the country . . .
Makranis, the Negroes of West Pakistan John B. Edlefsen, Khalida Shah and Mohsin Farooq Phylon (1960-) Vol. 21, No. 2 (2nd Qtr., 1960), pp. 124-130 Published by: Clark Atlanta University DOI: 10.2307/274335 Stable URL: http://www.jstor.org/stable/274335 Page Count: 7
quote:However, as for M1, minor N North-African influences have been detected by the presence of an U6 lineage in our Saudi sample. It has been suggested that the rare U9 clade might be another interesting exception because it has been detected only in Pakistan [26], Ethiopia, and Yemen [19], and now in our Saudi sample. U9 occurs frequently only among the Makrani population in Pakistan, which is characterized by a large component of sub-Saharan African lineages, suggesting that U9 lineages in Pakistan might also have an African origin [19]. Makrani sub-Saharan Africa lineages have exact matches in Africa, which is compatible with a recent conection as the result of the East African slave trade [26]. However, the entire sequenced Ethiopian and Pakistani U9 lineages [37] are separated by a mean of 4.5 coding mutations from the common root, placing the split at Paleolithic times. Most probably, Ethiopia received its U9 lineages from the Arabian Peninsula that, in turn, received them from northern areas. The southern geographic distribution of U9 contrasts with the west-northern distribution U4, of its sister clade [52], but this is a pattern shared with other Paleolithic U radiations such as U2, U7 [32], or U8 [53] that have eastern and western branches. [54].
--Kivisild et al
quote:"A potential issue that could in theory influence our findings is that the exact population contributing to African ancestry in West Eurasians is unknown. To gain insight into the African source populations, we carried out PCA analyses, which suggested that the African ancestry in West Eurasians is at least as closely related to East Africans (e.g. Hapmap3 Luhya (LWK)) as to West Africans (e.g. Nigerian Yoruba (YRI)) (the same analyses show that there is no evidence of relatedness to Chadic populations like Bulala) (Text S5 and Figure S12).
We also used the 4 Population Test to assess whether the tree ((LWK, YRI),(West Eurasian, CEU)) is consistent with the data, and found no evidence for a violation,
which is consistent with a mixture of either West African or East African ancestors or both contributing to the African ancestry in West Eurasians (Table S14; Figure S13). Historically, a mixture of West and East African ancestry is plausible, since African gene flow into West Eurasia is documented from both West Africa during Roman times [34] and from East Africa during migrations from Egypt [7]. It is important to point out, however, that the difficulty of pinpointing the exact African source population is not expected to bias our inferences about the total proportion and date of mixture. The f4 Ancestry Estimation method is unbiased even when we use a poor surrogates for the true ancestral African population (as long as the phylogeny is correct), as we confirmed by repeating analyses replacing YRI with LWK, and obtaining similar results (Table S15).Our ROLLOFF admixture date estimates are also similar whether we use LWK or YRI to represent ancestral African population (Table S15), as predicted by the theory.
--Moorjani et al.
quote:HAPLOGROUP L2A1
Haplogroup L2a1 was found in two specimens from the Southern Levant Pre-Pottery Neolithic B site at Tell Halula, Syria, dating from the period between ca. 9600 and ca. 8000 BP or 7500 - 6000 BCE.[13]
quote: "These results indicate that the ancestor of all Semitic languages in our dataset was being spoken in the Near East no earlier than approximately 7400 YBP, after having after having diverged from Afroasiatic in Africa"
(i) Semitic had an Early Bronze Age origin (approx. 5750 YBP) in the Levant, followed by an expansion of Akkadian into Mesopotamia;
(ii) Central and South Semitic diverged earlier than previously thought throughout the Levant during the Early to Middle Bronze Age transition; and
(iii) Ethiosemitic arose as the result of a single, possibly pre-Aksumite, introduction of a lineage from southern Arabia to the Horn of Africa approximately 2800 YBP.
Genomic Diversity: Applications in Human Population Genetics (1999, 2012)
--Luca Cavalli-Sforza
Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by Swenet: @Oshun
Sound analysis.
It should be noted, though, that R-V88, being a Y chromosome, is inherited as a single independent unit (meaning, independent from parts of the genome where most of our genetic heritage resides). Because of this, haplogroups don't have to have the affinity baggage associated with closely related haplogroups. So, even though R-V88 is derived from R, a Eurasian haplogroup, we don't know what the R-V88 people flowing into Africa were like, genetically.
In exactly the same way, R-V88 is today associated with Sub-Saharan African versions of sicklemia in some areas:
^So, even IF they find R-V88 in Nile Valley ancient DNA, it doesn't mean it belongs to someone who can be meaningfully called 'Eurasian' or 'non-African'.
Historically the Siwas Berbers are related to other Berbers with R-V88 from more Southern parts, like south Sahara.
quote: The remarkable archaeological site, dating back 10,000 years and called Gobero after the Tuareg name for the area, was brimming with skeletons of humans and animals including large fish and crocodiles. Gobero is hidden away within Nigers forbidding Ténéré Desert, known to Tuareg nomads as a desert within a desert.
Historically the Siwas Berbers are related to other Berbers with R-V88 from more Southern parts, like south Sahara.
Notice how the chart shows the exact opposite of what Ish Gebor is saying. In fact the Siwa show significant differences from other berbers, they have near no M81 (E1b1b1b) which is primary to the Y DNA of many other berbers and and the same time carry B-M109 ( aka B2a1a) which the other berbers on the chart carry none at all
Additionally it makes no sense to talk about V88 posting a chart with data compiled in 2002-2007 before the announcement of discovery of V88 in 2010 !!
And as per historically:
quote:
Th edifferentiationobservedbetweenNorth-WesternandNorth-EasternAf- rican Berbersseem sto be correlated w ith linguistic studiesw hich show that Siw an has a num ber of features w hich are only encountered in this oasis,and not else- w here in Berber...
First,in addition to the geo- graphicaldistance,that certainly increases the genetic distance,the geographical location ofBerber populations is very peculiar:the Berbers from the M aghreb are attheendofalongmigrationroute,whereasBerbersfrom Siwaareratherina crossroads betw een the M iddle East,East A frica,sub-Saharan areas and the N orth- A frican corridor.M eetings and exchanges betw een localand m igrating popula- tions w ere certainly not identical in N orth-W estern and N orth-E astern A frica. Then,the prehistory and history ofthe populations from the M aghreb are different from thoseoftheEgyptiangroup.
--- The Berber and the Berbers Genetic and linguistic diversities Jean-Michel Dugoujon1, Clotilde Coudray1, Antonio Torroni2, Fulvio Cruciani3, Rosaria Scozzari3, Pedro Moral4, Naima Louali5 and Maarten Kossmann6
2009
So even though Ish Gebor has a lot of quotes which he uses to avoid the topic article his analysis of the quotes is often wrong on more than one level, genetically and historically.
Again if you want to critique the thread topic than quite the article in the initial post. Simply posting other article quotes by themselves is trolling
Posted by Ish Gebor (Member # 18264) on :
LOL @ The above. How dumb. lol You absolutely have no knowledge on ethnography and the interaction of these ethic groups on this topic. Clown smh
According to lioness Africans were sitting on their "ape asses" doing nothing and had no abilities.
Notice how lioness is complete absent in understanding what was posted. It's laughable at best. lol
quote: Large-scale climate change forms the backdrop to the beginnings of food production in northeastern Africa (Kröpelin et al. 2008).[ Hunter-gatherer communities deserted most of the northern interior of the continent during the arid glacial maximum and took refuge along the North African coast, the Nile Valley, and the southern fringes of the Sahara (Barich and Garcea 2008; Garcea 2006; Kuper and Kröpelin 2006). During the subsequent Early Holocene African humid phase, from the mid-eleventh to the early ninth millennium cal BP, ceramic-using hunter-gatherers took advantage of more favorable savanna conditions to resettle much of northeastern Africa (Holl 2005; Kuper and Kröpelin 2006). Evidence of domestic animals first appeared in sites in the Western Desert of Egypt, the Khartoum region of the Nile, northern Niger, the Acacus Mountains of Libya, and Wadi Howar (Garcea 2004, 2006; Pöllath and Peters 2007; fig. 1).
--Fiona Marshall
Domestication Processes and Morphological Change Through the Lens of the Donkey and African Pastoralism Fiona Marshall and Lior Weissbrod
quote:Evidence from throughout the Sahara indicates that the region experienced a cool, dry and windy climate during the last glacial period, followed by a wetter climate with the onset of the current interglacial, with humid conditions being fully established by around 10,000 years BP, when we see the first evidence of a reoccupation of parts of the central Sahara by hunter gathers, most likely originating from sub-Saharan Africa (Cremaschi and Di Lernia, 1998; Goudie, 1992; Phillipson, 1993; Ritchie, 1994; Roberts, 1998).
[...]
Conical tumuli, platform burials and a V-type monument represent structures similar to those found in other Saharan regions and associated with human burials, appearing in sixth millennium BP onwards in northeast Niger and southwest Libya (Sivilli, 2002). In the latter area a shift in emphasis from faunal to human burials, complete by the early fifth millennium BP, has been interpreted by Di Lernia and Manzi (2002) as being associated with a changes in social organisation that occurred at a time of increasing aridity. While further research is required in order to place the funerary monuments of Western Sahara in their chronological context, we can postulate a similar process as a hypothesis to be tested, based on the high density of burial sites recorded in the 2002 survey. Fig. 2: Megaliths associated with tumulus burial (to right of frame), north of Tifariti (Fig. 1). A monument consisting of sixty five stelae was also of great interest; precise alignments north and east, a division of the area covered into separate units, and a deliberate scattering of quartzite inside the structure, are suggestive of an astronomical function associated with funerary rituals. Stelae are also associated with a number of burial sites, again suggesting dual funerary and astronomical functions (Figure 2). Further similarities with other Saharan regions are evident in the rock art recorded in the study area, although local stylistic developments are also apparent. Carvings of wild fauna at the site of Sluguilla resemble the Tazina style found in Algeria, Libya and Morocco (Pichler and Rodrigue, 2003), although examples of elephant and rhinoceros in a naturalistic style reminiscent of engravings from the central Sahara believed to date from the early Holocene are also present.
--Nick Brooks et al.
The prehistory of Western Sahara in a regional context: the archaeology of the "free zone"
Tyndall Centre for Climate Change Research, Saharan Studies Programme and School of Environmental Sciences, University of East Anglia, Norwich, UK Coauthors: Di Lernia, Savino ((Department of Scienze Storiche, Archeologiche, e Antropologiche dellAntichità, Faculty of Human Sciences, University of Rome La Sapienza, Via Palestro 63, 00185 Rome, Italy) and Drake, Nick (Department of Geography, Kings College, Strand, London WC2R 2LS).
quote: Large-scale climate change forms the backdrop to the beginnings of food production in northeastern Africa (Kröpelin et al. 2008).[ Hunter-gatherer communities deserted most of the northern interior of the continent during the arid glacial maximum and took refuge along the North African coast, the Nile Valley, and the southern fringes of the Sahara (Barich and Garcea 2008; Garcea 2006; Kuper and Kröpelin 2006). During the subsequent Early Holocene African humid phase, from the mid-eleventh to the early ninth millennium cal BP, ceramic-using hunter-gatherers took advantage of more favorable savanna conditions to resettle much of northeastern Africa (Holl 2005; Kuper and Kröpelin 2006). Evidence of domestic animals first appeared in sites in the Western Desert of Egypt, the Khartoum region of the Nile, northern Niger, the Acacus Mountains of Libya, and Wadi Howar (Garcea 2004, 2006; Pöllath and Peters 2007; fig. 1).
--Fiona Marshall
Domestication Processes and Morphological Change Through the Lens of the Donkey and African Pastoralism Fiona Marshall and Lior Weissbrod
Posted by Ish Gebor (Member # 18264) on :
According to lioness Africans had no interactions with one another, until eurasian came along to spread "their wisdom". lol From the posts by this dude we know that he advocates cacasoid theories, with "whiteness" as supreme ruling, over others. lol
Topology Atlas || Conferences
"Rapid and catastrophic environmental changes in the Holocene and human response" first joint meeting of IGCP 490 and ICSU Environmental catastrophes in Mauritania, the desert and the coast January 4-18, 2004
Field conference departing from Atar Atar, Mauritania
Organizers Suzanne Leroy, Aziz Ballouche, Mohamed Salem Ould Sabar, and Sylvain Philip (Hommes et Montagnes travel agency)
View Abstracts Conference Homepage
What is the impact of Holocene climatic changes on human societies: analysis of Neolithic population dynamic and dietary customs. by Jousse, Helene
UMR Paléoenvironnements et Paléobiosphère, Université Claude Bernard Lyon 1, Villeurbanne, France.
quote: The reconstruction of human cultural patterns in relation to environmental variations is an essential topic in modern archaeology.
In western Africa, a first Holocene humid phase beginning c. 11,000 years BP is known from the analysis of lacustrine sediments (Riser, 1983 ; Gasse, 2002). The monsoon activity increased and reloaded hydrological networks (like the Saharan depressions) leading to the formation of large palaeolakes. The colonisation of the Sahara by vegetation, animals and humans was then possible essentially around the topographic features like Ahaggar (fig. 1). But since 8,000 years BP, the climate began to oscillate towards a new arid episode, and disturbed the ecosystems (Jolly et al., 1998; Jousse, 2003).
First, the early Neolithics exploited the wild faunas, by hunting and fishing, and occupied small sites without any trace of settlement in relatively high latitudes. Then, due to the climatic deterioration, they had to move southwards.
This context leads us to consider the notion of refugia. Figure 1 presents the main zones colonised by humans in western Africa. When the fossil valleys of Azaouad, Tilemsi and Azaouagh became dry, after ca. 5,000 yr BP, humans had to find refuges in the Sahelian belt, and gathered around topographic features (like the Adrar des Iforas, and the Mauritanians Dhar) and major rivers, especially the Niger Interior Delta, called the Mema.
Whereas the Middle Neolithic is relatively well-known, the situation obviously becomes more complex and less information is available concerning local developments in late Neolithic times.. Only some cultural affiliations existed between the populations of Araouane and Kobadi in the Mema. Elsewhere, and especially along the Atlantic coast and in the Dhar Tichitt and Nema, the question of the origin of Neolithic peopling remains unsolved.
A study of the palaeoenvironment of those refugia was performed by analysing antelopes ecological requirements (Jousse, submitted). It shows that even if the general climate was drying from 5,000 4,000 yr BP in the Sahara and Sahel, edaphic particularities of these refugia allowed the persistence of local gallery forest or tree savannas, where humans and animals could have lived (fig. 2). At the same time, cultural innovation like agriculture, cattle breeding, social organisation in villages are recognised. For the moment, the relation between the northern and the southern populations are not well known.
How did humans react against aridity? Their dietary behaviour are followed along the Holocene, in relation with the environment, demographic expansion, settling process and emergence of productive activities.
- The first point concerns the pastoralism. The progression of cattle pastoralism from eastern Africa (fig. 3) is recorded from 7,400 yr BP in the Ahaggar and only from 4,400 yr BP in western Africa. This trend of breeding activities and human migrations can be related to climatic evolution. Since forests are infested by Tse-Tse flies preventing cattle breeding, the reduction of forest in the low-Sahelian belt freed new areas to be colonised. Because of the weakness of the archaeozoological material available, it is difficult to know what was the first pattern of cattle exploitation.
- A second analysis was carried on the resources balance, between fishing-hunting-breeding activities. The diagrams on figures 4 and 5 present the number of species of wild mammals, fishes and domestic stock, from a literature compilation. Fishing is known around Saharan lakes and in the Niger. Of course, it persisted with the presence of water points and even in historical times, fishing became a specialised activity among population living in the Niger Interior Delta. Despite the general environmental deterioration, hunting does not decrease thanks to the upholding of the vegetation in these refugia (fig. 2). On the contrary, it is locally more diversified, because at this local scale, the game diversity is closely related to the vegetation cover. Hence, the arrival of pastoral activities was not prevalent over other activities in late Neolithic, when diversifying resources appeared as an answer to the crisis.
This situation got worse in the beginning of historic times, from 2,000 yr BP, when intense settling process and an abrupt aridity event (Lézine & Casanova, 1989) led to a more important perturbation of wild animals communities. They progressively disappeared from the human diet, and the cattle, camel and caprin breeding prevailed as today.
Gasse, F., 2002. Diatom-inferred salinity and carbonate oxygen isotopes in Holocene waterbodies of the western Sahara and Sahel (Africa). Quaternary Science Reviews: 717-767.
Jolly, D., Harrison S. P., Damnati B. and Bonnefille R. , 1998. Simulated climate and biomes of Africa during the late Quaternary : Comparison with pollen and lake status data. Quaternary Science Review 17: 629-657.
Jousse H., 2003. Impact des variations environnementales sur la structure des communautés mammaliennes et l'anthropisation des milieux: exemple des faunes holocènes du Sahara occidental. Thèse de lUniversité Lyon 1, 405 p.
Jousse H, 2003. Using archaeological fauna to calibrate palaeovegetation: the Holocene Bovids of western Africa. Submit to Quaternary Science Reviews in november 2003, référence: QSR 03-333.
Lézine, A. M. and J. Casanova, 1989. Pollen and hydrological evidence for the interpretation of past climate in tropical West Africa during the Holocene. Quaternary Science Review 8: 45-55.
Riser, J., 1983. Les phases lacustres holocènes. Sahara ou Sahel ? Quaternaire récent du bassin de Taoudenni (Mali). Marseille: 65-86.
quote: The presence of sub-Saharan L-type mtDNA sequences in North Africa has traditionally been explained by the recent slave trade. However, gene flow between sub-Saharan and northern African populations would also have been made possible earlier through the greening of the Sahara resulting from Early Holocene climatic improvement. In this article, we examine human dispersals across the Sahara through the analysis of the sub-Saharan mtDNA haplogroup L3e5, which is not only commonly found in the Lake Chad Basin (∼17%), but which also attains nonnegligible frequencies (∼10%) in some Northwestern African populations. Age estimates point to its origin ∼10 ka, probably directly in the Lake Chad Basin, where the clade occurs across linguistic boundaries. The virtual absence of this specific haplogroup in Daza from Northern Chad and all West African populations suggests that its migration took place elsewhere, perhaps through Northern Niger. Interestingly, independent confirmation of Early Holocene contacts between North Africa and the Lake Chad Basin have been provided by craniofacial data from Central Niger, supporting our suggestion that the Early Holocene offered a suitable climatic window for genetic exchanges between North and sub-Saharan Africa. In view of its younger founder age in North Africa, the discontinuous distribution of L3e5 was probably caused by the Middle Holocene re-expansion of the Sahara desert, disrupting the clade's original continuous spread.
--Elika Podgorná et al.
Annals of Human Genetics Volume 77, Issue 6, pages 513523, November 2013
The Genetic Impact of the Lake Chad Basin Population in North Africa as Documented by Mitochondrial Diversity and Internal Variation of the L3e5 Haplogroup
Historically the Siwas Berbers are related to other Berbers with R-V88 from more Southern parts, like south Sahara.
Notice how the chart shows the exact opposite of what Ish Gebor is saying. In fact the Siwa show significant differences from other berbers, they have near no M81 (E1b1b1b) which is primary to the Y DNA of many other berbers and and the same time carry B-M109 ( aka B2a1a) which the other berbers on the chart carry none at all
Do you not see how you are debunking yourself. lol smh
You will dispute R-V88 being an old reference in this chard, but you will refer to M81 and B-M109, which I didn't not even mention. lol You hypocrite.
But B-M109 relates to the people of the region we speak of here who carry R-V88. lol So you shot yourself in your eurocentric foot again. You need to lie some more. Back to the drawing table for you, make up another lie to cover up a lie.
The 1st PC reflects an axial proportion of E3a* vs. E1* where Papel and Felupe-Djola retain the higher proportions of the later. E3a* is again a main influence in the 2ndaxis against that of R1b and E3b1 Posted by the lioness, (Member # 17353) on :
quote:Originally posted by Ish Gebor:
quote:Originally posted by the lioness,:
quote:Originally posted by Ish Gebor:
Historically the Siwas Berbers are related to other Berbers with R-V88 from more Southern parts, like south Sahara.
Notice how the chart shows the exact opposite of what Ish Gebor is saying. In fact the Siwa show significant differences from other berbers, they have near no M81 (E1b1b1b) which is primary to the Y DNA of many other berbers and and the same time carry B-M109 ( aka B2a1a) which the other berbers on the chart carry none at all
Do you not see how you are debunking yourself. lol smh
You will dispute R-V88 being an old reference in this chard, but you will refer to M81 and B-M109, which I didn't not even mention. lol You hypocrite.
But B-M109 relates to the people of the region we speak of here who carry R-V88. lol So you shot yourself in your eurocentric foot again. You need to lie some more. Back to the drawing table for you, make up another lie to cover up a lie.
You continue not knowing how to interpret the data properly.
Again, B-M109 relates to the people of Cameroon where V88 is also present yet you fail to mention that only the Siwa carry it - an example of them not relating to other berbers.
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by the lioness,:
quote:Originally posted by Ish Gebor:
quote:Originally posted by the lioness,:
quote:Originally posted by Ish Gebor:
Historically the Siwas Berbers are related to other Berbers with R-V88 from more Southern parts, like south Sahara.
Notice how the chart shows the exact opposite of what Ish Gebor is saying. In fact the Siwa show significant differences from other berbers, they have near no M81 (E1b1b1b) which is primary to the Y DNA of many other berbers and and the same time carry B-M109 ( aka B2a1a) which the other berbers on the chart carry none at all
Do you not see how you are debunking yourself. lol smh
You will dispute R-V88 being an old reference in this chard, but you will refer to M81 and B-M109, which I didn't not even mention. lol You hypocrite.
But B-M109 relates to the people of the region we speak of here who carry R-V88. lol So you shot yourself in your eurocentric foot again. You need to lie some more. Back to the drawing table for you, make up another lie to cover up a lie.
You continue not knowing how to interpret the data properly.
Again, B-M109 relates to the people of Cameroon where V88 is also present yet you fail to mention that only the Siwa carry it - an example of them not relating to other berbers.
You are a DUMB BOX OF ROCKS!!!! There is no need to reason with you on logic, since you lack that ability. And the major problem here is that you try to speak on people and ethnic groups you have literarily no understand about. But for some odd reason you think you are bright. lol But in realty you're a dishonest euronut.
quote: The Tuareg presently live in the Sahara and the Sahel. Their ancestors are commonly believed to be the Garamantes of the Libyan Fezzan, ever since it was suggested by authors of antiquity. Biological evidence, based on classical genetic markers, however, indicates kinship with the Beja of Eastern Sudan. Our study of mitochondrial DNA (mtDNA) sequences and Y chromosome SNPs of three different southern Tuareg groups from Mali, Burkina Faso and the Republic of Niger reveals a West Eurasian-North African composition of their gene pool. The data show that certain genetic lineages could not have been introduced into this population earlier than ∼9000 years ago whereas local expansions establish a minimal date at around 3000 years ago.
--Luísa Pereira, et al.
Eur J Hum Genet. 2010 Aug; 18(8): 915923. Published online 2010 Mar 17. doi: 10.1038/ejhg.2010.21
quote:This site has been called Gobero, after the local Tuareg name for the area. About 10,000 years ago (77006200 B.C.E.), Gobero was a much less arid environment than it is now. In fact, it was actually a rather humid lake side hometown of sorts for a group of hunter-fisher-gatherers who not only lived their but also buried their dead there. How do we know they were fishing? Well, remains of large nile perch and harpoons were found dating to this time period.
BTW, the above is what they claimed as "their" suppose backmigration, we've seen earlier on. They try to claim it as theirs. Which is ridiculous of course.
lol As I stated from the beginning Siwa Berbers are close to Berbers from Southern parts such as, south of the Sahara. You fail to use your brains.
Posted by the lioness, (Member # 17353) on :
quote:Originally posted by Ish Gebor:
Historically the Siwas Berbers are related to other Berbers with R-V88 from more Southern parts, like south Sahara.
^^ He doesn't know what he's talking about
In the Maghreb countries, the frequency of R-V88 drops to around 1%.
--Maybe a whole bunch of copy and paste quotes can give the illusion that he knows what he's talking about, yet we know what's up
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by the lioness,:
quote:Originally posted by Ish Gebor:
Historically the Siwas Berbers are related to other Berbers with R-V88 from more Southern parts, like south Sahara.
^^ He doesn't know what he's talking about
In the Maghreb countries, the frequency of R-V88 drops to around 1%.
--Maybe a whole bunch of copy and paste quotes can give the illusion that he knows what he's talking about, yet we know what's up
Sweet Lord, why? Why?
Here, this is more on your level,
"Berbers in Niger are Niger citizens of Berber descent or persons of Berber descent residing in Niger. Ethnic Berbers in Niger are believed to number of 1,620,000[2]"
quote: The Gobero Archaeological Project is a partnership between Arizona State University and the University of Chicago that involves over a dozen international collaborators across the fields of geology, archaeology, chemistry, and paleontology. The research focuses on a series of Early and Middle Holocene cemeteries (circa 10,000-5000 years ago) from the southern Sahara Desert (Niger) where a unique record of human life and death was preserved. The project combines ongoing fieldwork with extensive archaeological and bioarcheological lab analyses of human and non-human skeletal and dental remains, ceramics, lithics and soils to reconstruct the sequence of human occupations in a region that is now desolate and uninhabited.
In addition, we seek to reconstruct the nature of early Saharan communities in terms of their mortuary practices, social organization, life courses and health experiences. A key component of the work is evaluating the transition to food production where, in North Africa, the Neolithic model is reversed: populations become more mobile and adopted cattle and caprine pastoralism well before agriculture. The projects results are contextualized within the wider Saharan interaction sphere and linked to ongoing climatic reconstructions of the Holocene Sahara.
^ He puts up endless maps and quotes as a reply and they are not a reply, just endless diversion
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by the lioness,: ^ He puts up endless maps and quotes as a reply and they are not a reply, just endless diversion
LOL Lioness, you're dumb and don't know the ethnic groups. That is the conclusion here. And above all, you're a dishonest euronut.Typical.
quote: The language and culture of the Berbers are seeing a popular resurgence. However, the nationalism of some Tuareg groups (in Mali and northern Niger) threatens the peace of the region.
quote:Among the Niger-Congo-speaking populations, the frequency of the haplogroup R-V88 ranged between 0.0 and 66.7%. Outside central Africa, haplogroup R-V88 was only observed in Afroasiatic-speaking populations from northern Africa, with frequencies ranging from 0.3% in Morocco, to 3.0% in Algeria, and to 11.5% in Egypt, where a particularly high frequency (26.9%) was observed among the Berbers from the Siwa Oasis.
--Fulvio Cruciani et al.
Human Y chromosome haplogroup R-V88: a paternal genetic record of early mid Holocene trans-Saharan connections and the spread of Chadic languages
European Journal of Human Genetics (2010) 18, 800807; doi:10.1038/ejhg.2009.231; published online 6 January 2010
quote: The Tuareg are seminomadic pastoralist groups that are mostly spread between Libya, Algeria, Mali, and Niger. They speak a Berber language and are believed to be the descendents of the Garamantes people of Fezzan, Libya (500 BC - 700 CE) [34].
--Karima Fadhlaoui-Zid et al.
Genome-Wide and Paternal Diversity Reveal a Recent Origin of Human Populations in North Africa
November 27, 2013http://dx.doi.org/10.1371/journal.pone.0080293
But you know what lioness, I am buzzy with other things such as software development, so I don't post no longer as often. This was actually waste of my time, trying to explain to you something this simple.
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by Ish Gebor:
quote:Originally posted by the lioness,:
quote:
The Ain Dokhara specimen also falls within the scatter of recent African means. In contrast, none of the Afalou specimens, nor of the El Wad Natufian specimens, falls within the African scatter, and all lie toward the more cold-adapted end of the scatter.
Conclusions
....The Jebel Sahaba sample are distinct in body shape from penecontemporary humans from Afalou-Bou- Rhummel (Algeria) and El Wad Natufians from the southern Levanta result consistent with the results of both Irish (2000, 2005) using dental data and Franciscus (1995, 2003) using nasal data.
--2013 DOI: 10.1002/oa.2315 Population Affinities of the Jebel Sahaba Skeletal Sample: Limb Proportion Evidence T. W. HOLLIDAY*
Interesting is:
quote: These results corroborate those of both Irish and Franciscus, who, using dental, oral and nasal morphology,[...]
The body shape of the terminal Pleistocene Jebel Sahaba population is tropical-adapted, with elongated limbs, especially in the distal segments, and is most similar to living sub-Saharan Africans and less similar to late Pleistocene and Holocene North Africans (including Egyptians and Nubians).[...]
Of which sub-Saharan Africans do that speak?
Btw,
quote: As with the previous analysis, the North Africans are intermediate between the sub- Saharan Africans and the Europeans, whereas the Europeans tend toward longer tibiae than the Inuits.
--T. W. HOLLIDAY
Population Affinities of the Jebel Sahaba Skeletal Sample: Limb Proportion Evidence
Posted by Djehuti (Member # 6698) on :
Again, I ask why it seems the author is trying to associate R1 in Africa with Chadic or Afrasian when it's highest frequency is among Niger-Congo speakers?
Also, why associate R1 (R1b or otherwise) with Afrasian or any language phylum at all when it predates all modern language phyla.
The irony is that there are far more Afrasian speakers in Africa who carry the Eurasian clade hg T than R!
Posted by the lioness, (Member # 17353) on :
quote:Originally posted by Djehuti: [QB] Again, I ask why it seems the author is trying to associate R1 in Africa with Chadic or Afrasian when it's highest frequency is among Niger-Congo speakers?
The Fang people are relatively recent migrants into the Equatorial Guinea, and many of them moved from central Cameroon in 19th century.
Posted by Djehuti (Member # 6698) on :
^ That still doesn't answer my question. Are you saying Cameroonians are not indigenous but Eurasian?
Posted by the lioness, (Member # 17353) on :
quote:Originally posted by the lioness,: [QB] [QUOTE]Originally posted by Djehuti: [QB] Again, I ask why it seems the author is trying to associate R1 in Africa with Chadic or Afrasian when it's highest frequency is among Niger-Congo speakers?
quote:
[QUOTE]
Within Africa, the highest frequencies of the R-V88 haplogroup (and its commonest sub-clade, R-V69) were observed in the central Sahel (northern Cameroon, northern Nigeria, Chad, and Niger) (Table 1; Figure 3). Immediately south of this region (southern Cameroon and southern Nigeria), frequencies drastically dropped to 0.04.8%. The central Sahel is characterized by a strong linguistic fragmentation with populations speaking languages belonging to three of the four linguistic families of Africa (Afroasiatic, Niger-Congo, and Nilo-Saharan). When the linguistic affiliation of the populations from the central Sahel was also taken into account, a clear-cut divide was observed between those speaking Afroasiatic languages (including the Berber-speaking Tuareg, the Semitic Arab Shuwa, and Chadic-speaking populations from northern Cameroon) and the other populations (MannWhitney test P=1.4 × 10−3), with Chadic-speaking populations mostly contributing to this difference. It is worth noting that, if the finding of 20% R-V88 chromosomes among the Hausa (Table 1) is representative, this population, encompassing by far more people than all other Chadic speakers,44 also encompasses the highest absolute number of V88 carriers.
In contrast to prior studies on nuclear (mostly autosomal) ins/del and microsatellite markers,45 the Chadic are distinguished from the Nilo-Saharan-speaking populations at the Y chromosome variation level (Table 1; Supplementary Table 3). Repeated assimilations of Nilo-Saharan females over generations may account for these conflicting signals. Among the Niger-Congo-speaking populations, the frequency of the haplogroup R-V88 ranged between 0.0 and 66.7%. Outside central Africa, haplogroup R-V88 was only observed in Afroasiatic-speaking populations from northern Africa, with frequencies ranging from 0.3% in Morocco, to 3.0% in Algeria, and to 11.5% in Egypt, where a particularly high frequency (26.9%) was observed among the Berbers from the Siwa Oasis. Although the presence of the haplogroup R-V88 at non-negligible frequencies in some Niger-Congo-speaking populations from the central Sahel can be accounted for by Chadic admixture favored by geographic contiguity, the presence of this haplogroup both in northern Africa and the central Sahel is especially intriguing given that >1500 km across the Sahara separate the two regions. The expansion time for the haplogroup R-V88 in Africa, under two different population models (see Materials and methods), was found to be 9.25.6 ky (95% CI=7.610.8 ky and 4.76.6 ky, respectively).
Diverse hypotheses have been proposed to explain the process by which proto-Chadic speakers arrived to the Lake Chad region. Ehret46 has put forward a model for Afroasiatic languages with a primary division between the Omotic languages of Ethiopia and an Erythraean subgroup. This, in turn, has been subdivided into Cushitic and North Erythraean, the latter including Berber, Semitic, Ancient Egyptian, and Chadic. In his opinion, around 7000 kya proto-Chadic Afroasiatic speakers may have moved west through the Central Sahara and then farther south into the Lake Chad Basin.47 Blench,48 in turn, suggested that speakers of proto-CushiticChadic language migrated east-to-west from the Middle Nile to the Lake Chad, and recent mtDNA data support this view.49 However, in contrast to the mtDNA, a strong connection between Chadic and other Afroasiatic populations from Northern Africa is revealed by the Y chromosome data. This finding would indicate the trans-Saharan47 a more likely scenario than the inter-Saharan hypothesis,48 at least as far as the male component of gene pool is concerned. In this view, it is tempting to speculate that the Y chromosome haplogroup R-V88 represents a preserved genetic record of gene flow along the same axis as the proposed spread of proto-Chadic languages.47 Indeed, geomorphological evidence4 from the paleolakes that existed in the Sahara during the mid-Holocene indicates that these lakes could have covered an area as large as about 10% of the Sahara, providing an important corridor for human migrations across the region.5
In summary, our data indicate a significant male contribution from northern Africa (and ultimately Asia) to the gene pool of the central Sahel. The trans-Saharan population movements resulting in this genetic pattern would seem to mirror the spread of the proto-Chadic languages, and most likely took place during the early mid Holocene, a period when giant paleolakes may have provided a corridor for human migrations across what is now the Sahara desert.
Human Y chromosome haplogroup R-V88: a paternal genetic record of early mid Holocene trans-Saharan connections and the spread of Chadic languages
Fulvio Cruciani 2010
^ see Ish Gebor, this is how to quote properly. The quote is directly related to answer the question rather than on a tangent
Posted by Elmaestro (Member # 22566) on :
quote:Originally posted by Djehuti: ^ That still doesn't answer my question. Are you saying Cameroonians are not indigenous but Eurasian?
Smokescreen, and their occams razor lol It was 2016 when this paper was published - probably 2015 when the study was completed. Thou with any updated historical knowledge knows V88 isn't an A-Asiatic indicator. However wording that the possible Eurasian backflow via V88 contributed to the spread of AA in SSA nudges readers to believe Afroasiatic and other Sahelian cultural phenomenon are not a product of SSAn development.
Lets not focus on the fact that we have V88 possibly existing almost 10k years before it's emergence in Chad-Cameroon according to this paper
quote:We estimate that the Eurasian R1b lineages initially diverged 7,3009,400 ya, at the time of the Neolithic expansions. However, we found that the African and Eurasian R1b lineages diverged 17,90023,000 ya, suggesting that genetic structure was already established between the groups who expanded to Europe and Africa.
...Ok So this means 1 of two things, The so called African Haplogroup of v88 (non derived) should be detectable OOA or Precursor Haplogroups should be found somewhere around chad or Cameroon because...
quote: We estimate that mixture occurred 4,7507,200 ya, thus after the Neolithic transition in the Near East, a period characterized by exponential growth in human population size.
Due to the fact that they dated the Chadian R1b branch to before the date directly above We should expect to see some chadian R1b well outside of chad, The simple fact that we have V88(derived) possibly emerging within the time frame of admixture is fucking convenient either way you put it.
quote:tip.We estimate that the Chadian R1b emerged 5,7007,300 ya, whereas most European R1b haplogroups emerged 7,3009,400 ya.
DOI:10.1016/j.ajhg.2016.10.012 Posted by the lioness, (Member # 17353) on :
Posted by the lioness, (Member # 17353) on :
there is some interesting commentary of the thread topic article here;
quote:Originally posted by Djehuti: Again, I ask why it seems the author is trying to associate R1 in Africa with Chadic or Afrasian when it's highest frequency is among Niger-Congo speakers?
Also, why associate R1 (R1b or otherwise) with Afrasian or any language phylum at all when it predates all modern language phyla.
The irony is that there are far more Afrasian speakers in Africa who carry the Eurasian clade hg T than R!
It's because Chadic speaking groups carry that marker, all over West Africa. And secondly to explain civilization as intrusion.
I remember this earlier discussion on HgT, perhaps it is useful.
quote:Originally posted by the lioness,: ^ see Ish Gebor, this is how to quote properly. The quote is directly related to answer the question rather than on a tangent
This branching pattern, along with the geographical distribution of the major clades A, B, and CT, has been interpreted as supporting an African origin for anatomically modern humans,10 with Khoisan from south Africa and Ethiopians from east Africa sharing the deepest lineages of the phylogeny.15 and 16
[...]
The deepest branching separates A1b from a monophyletic clade whose members (A1a, A2, A3, B, C, and R) all share seven mutually reinforcing derived mutations (five transitions and two transversions, all at non-CpG sites).
[...]
How does the present MSY tree compare with the backbone of the recently published reference MSY phylogeny?13 The phylogenetic relationships we observed among chromosomes belonging to haplogroups B, C, and R are reminiscent of those reported in the tree by Karafet et al.13 These chromosomes belong to a clade (haplogroup BT) in which chromosomes C and R share a common ancestor (Figure 2).
quote:The haplogroup R1b1a (R-V88) was found with a fre- quency of 8% in the village of Al Awaynat. Generally, haplogroup R and its subsets are spread in Eurasia as far as Siberia (Karafet et al., 2008; Chiaroni et al., 2009; Lancaster, 2010). Nevertheless, R1b1a has been observed at high frequencies in Northwest Africa (27% in the Egyptian Berbers), with peaks in the Chadic-speaking populations from Central Africa, ranging from 29 to 96% in Cameroon, and very rarely is found outside Africa (Cruciani et al., 2010a,b). This haplogroup has been pro- posed to represent the paternal genetic signature of the mid-Holocene migration of proto-Chadic Afro-asiatic speakers across the Central Sahara to Lake Chad (Ehret, 2002; Cruciani et al., 2010a); this suggests a link between Chadic speakers and other Afro-Asiatic speak- ers to the north of the Sahara. In the eight-microsatellite Network analysis of R1b1a chromosomes from Northern and Central Africa (Fig. S2), the Libyan Tuareg R1b1a Y-chromosomes were found to belong to a branch characterized exclusively by haplotypes from Central Africa, more particularly from the Chad area (Cruciani et al., 2010a). This may be likely explained by recent introduction through the slavery practices mentioned above. Nonetheless, the hypothesis that the Libyan Tuareg R1b1a haplotypes may be relics of the migration of Pastoral proto-Chadic speakers, as hypothesized by the trans-Saharan hypothesis (Ehret, 2002; Cruciani et al., 2010a), cannot be ruled out
--Claudio Ottoni, et al. (2011)
Deep Into the Roots of the Libyan Tuareg: A Genetic Survey of Their Paternal Heritage
Posted by Djehuti (Member # 6698) on :
Sorry, here is another image of hg T distribution.
This topic was covered numerous times including here, here, and here
Not surprisingly Eurocentrists like Lioness seem to be fixated on alleged Eurasian clades like R which are relatively rare overall in the African continent while ignoring the blatant E clade in their homeland of Europe which accounts for one-third of the Y chromosomes of that subcontinent!
Posted by Elmaestro (Member # 22566) on :
^Didn't you check out the link to eupedia Lioness posted?? it seems there are people out there whom believe Hg E Is in fact Eurasian as well!! lol... though She/he/it was right, there were some interesting commentary on this article.
Posted by Doug M (Member # 7650) on :
Posting this again.
quote:Originally posted by Doug M: Here is the key question that people need to be asking and I know this has been discussed here before but it needs to be reiterated. How come African genes outside Africa are quickly labeled as something other than African because of splits, yet so-called Eurasian genes in Africa are Eurasian even thousands of years later and after multiple splits?
See the pattern here? Multiple papers in a short time period talking of ancient "Eurasian" lineages in Africa, yet Africa is the BASIS of all humans on the planet and there are HARDLY ANY papers on African lineages in Europe, the Levant, Asia and elsewhere. Sounds like a double standard to me.
The following wiki page shows how arbitrary and contradictory the logic goes regarding labeling African genetics versus Eurasian genetics. By the logic they use to label "Eurasian" mixture in Africa, ALL HUMANS outside of Africa and their lineages should be labeled as African for the first 10,000 years of human existence outside of Africa. But note that they don't do this.... Wonder why? Keep in mind that Neanderthal admixture is one scam they use to justify this nonsense.....
Haplogroup R-V88 is thought to originated in thew Cameroon/Chad region
Posted by Djehuti (Member # 6698) on :
quote:Originally posted by Elmaestro: ^Didn't you check out the link to eupedia Lioness posted?? it seems there are people out there whom believe Hg E Is in fact Eurasian as well!! lol... though She/he/it was right, there were some interesting commentary on this article.
Even mainstream geneticists have come to the consensus that hg E-M96 most likely arose in Africa especially since its ancestral clade DE* was found mainly in West African populations. Even if one were to go with the hypothesis that E arose in Eurasia, it would have risen somewhere in Southwest Asia around the the time of the first major OOA event and then immediately migrated right back to Africa where it expanded and diversified. LOL Even if one were to go by this more convoluted path, I doubt these back-migrating 'Eurasians' would have been any different from their African kin whom they only recently separated from only to rejoin!
Getting back to the topic, I fail to see how hg R (V88 or otherwise) in Chad has anything to do with Chadic language.
"It is possible from this overview of the data to conclude that the limited conceptual vocabulary shared by the ancestors of contemporary Chadic-speakers (therefore also contemporary Cushitic-speakers), contemporary Nilotic-speakers and Ancient Egyptian-speakers suggests that the earliest speakers of the Egyptian language could be located to the south of Upper Egypt (Diakonoff 1998) or, earlier, in the Sahara (Wendorf 2004), where Takács (1999, 47) suggests their long co-existence can be found. In addition, it is consistent with this view to suggest that the northern border of their homeland was further than the Wadi Howar proposed by Blench (1999, 2001), which is actually its southern border. Neither Chadics nor Cushitics existed at this time, but their ancestors lived in a homeland further north than the peripheral countries that they inhabited thereafter, to the south-west, in a Niger-Congo environment, and to the south-east, in a Nilo-Saharan environment, where they interacted and innovated in terms of language. From this perspective, the Upper Egyptian cultures were an ancient North East African periphery at the crossroads, as suggested by Dahl and Hjort-af-Ornas of the Beja (Dahl and Hjort-af-Ornas 2006). The most likely scenario could be this: some of these Saharo-Nubian populations spread southwards to Wadi Howar, Ennedi and Darfur; some stayed in the actual oases where they joined the inhabitants; and others moved towards the Nile, directed by two geographic obstacles, the western Great Sand Sea and the southern Rock Belt. Their slow perambulations led them from the area of Sprinkle Mountain (Gebel Uweinat) to the east Bir Sahara, Nabta Playa, Gebel Ramlah, and Nekhen/Hierakonpolis (Upper Egypt), and to the north-east by way of Dakhla Oasis to Abydos (Middle Egypt)."--Anselin (2009)
Posted by Swenet (Member # 17303) on :
quote:Originally posted by the lioness,: Haplogroup R-V88 is thought to originated in thew Cameroon/Chad region
The 7ky old V88 Y chromosome in Iberia casts doubt on V88 originating in West/Central Africa. It can't be in West/Central Africa and Iberia at the same time so close to its estimated age. You're better off stating that most or all of the current variation of R-V88 comes from the Cameroon/Chad region.
I think you might have missed that extra information because I see that you quoted an older version of that post at the time that doesn't contain the caveats I put in there.
Posted by Clyde Winters (Member # 10129) on :
Just because the oldest instance of a probable R1b-V88 in Iberia dates to 7,100ya (ID I0410, Els Trocs, Spain, Haak et al. 2015), does not make it older than the Chadian clade. Researchers have found that the TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010). Eurasians carry the M269 (R1b1b2) mutation. The subclades of R1b1b2 include Rh1b1b2g (U106) (TMRCA 8.3kya) and R1b1b2h (U152) (TMRCA 7.4kya). The most recent common ancestor for R1b1b2 is probably 8kya (Balaresque et al, 2010).
The fact that the earliest R1b aY-chromosome comes from Iberia should not be a surprise since West Africans frequently crossed into Eurasia via Iberia, instead of the Levant.
In Africa we find V45 (R-M207) and V88. Clearly, R-V88 in Chad is older than V88 in Iberia, Most importantly there is no archaeological evidence of a back migration of haplogroup R into Africa, but there is evidence of the migration of the Kushites and Proto-Saharans into Eurasia from Middle Africa. This supports the proposition the R haplogroups originated in Africa, not Eurasia.
Posted by Swenet (Member # 17303) on :
Reread what I said, including the linked posts.
Posted by Oshun (Member # 19740) on :
quote:Originally posted by Swenet:
quote:Originally posted by the lioness,: Haplogroup R-V88 is thought to originated in thew Cameroon/Chad region
The 7ky old V88 Y chromosome in Iberia casts doubt on V88 originating in West/Central Africa. It can't be in West/Central Africa and Iberia at the same time so close to its estimated age. You're better off stating that most or all of the current variation of R-V88 comes from the Cameroon/Chad region.
I think you might have missed that extra information because I see that you quoted an older version of that post at the time that doesn't contain the caveats I put in there.
But weren't Iberians around this time Africans?
2005 Eva Fernández Domínguez
Nerja, Málaga (Spain)
NE-NM82.2 (Sample Name 2NE): Haplogroup J*: 17000-20000 NE-NAP (Sample Name 4NE): Haplogroup L1b: 2,260 BC NE-1829 (Sample Name 5NE): Haplogroup L1b: 5.875±80 A.P.
Toledo, Portugal K-13 (Sample Name: TO1): Haplogroup L3a: 9200-7800 A.P.
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by Elmaestro: ^Didn't you check out the link to eupedia Lioness posted?? it seems there are people out there whom believe Hg E Is in fact Eurasian as well!! lol... though She/he/it was right, there were some interesting commentary on this article.
Yeah, that is what I stated earlier on. Lioness advocated this one as well on a few occasions.
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by Swenet:
quote:Originally posted by the lioness,: Haplogroup R-V88 is thought to originated in thew Cameroon/Chad region
The 7ky old V88 Y chromosome in Iberia casts doubt on V88 originating in West/Central Africa. It can't be in West/Central Africa and Iberia at the same time so close to its estimated age. You're better off stating that most or all of the current variation of R-V88 comes from the Cameroon/Chad region.
I think you might have missed that extra information because I see that you quoted an older version of that post at the time that doesn't contain the caveats I put in there.
Swenet,
What are the mtDNA L* markers correlating with these West / Central African populations?
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by Djehuti: Sorry, here is another image of hg T distribution.
This topic was covered numerous times including here, here, and here
Not surprisingly Eurocentrists like Lioness seem to be fixated on alleged Eurasian clades like R which are relatively rare overall in the African continent while ignoring the blatant E clade in their homeland of Europe which accounts for one-third of the Y chromosomes of that subcontinent!
Thanks for posting these.
I noticed The Explorer, posted 08 January, 2010 17:27. It is exactly what I have been posting on Fulvio Cruciani et al (2011), in the "A Revised Root for the Human Y Chromosomal Phylogenetic Tree: The Origin of Patrilineal Diversity in Africa".
But at EG it was figured out, prior to the 2011 paper, which is amazing. lol
quote:Originally posted by Clyde Winters: Just because the oldest instance of a probable R1b-V88 in Iberia dates to 7,100ya (ID I0410, Els Trocs, Spain, Haak et al. 2015), does not make it older than the Chadian clade. Researchers have found that the TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010). Eurasians carry the M269 (R1b1b2) mutation. The subclades of R1b1b2 include Rh1b1b2g (U106) (TMRCA 8.3kya) and R1b1b2h (U152) (TMRCA 7.4kya). The most recent common ancestor for R1b1b2 is probably 8kya (Balaresque et al, 2010).
The fact that the earliest R1b aY-chromosome comes from Iberia should not be a surprise since West Africans frequently crossed into Eurasia via Iberia, instead of the Levant.
In Africa we find V45 (R-M207) and V88. Clearly, R-V88 in Chad is older than V88 in Iberia, Most importantly there is no archaeological evidence of a back migration of haplogroup R into Africa, but there is evidence of the migration of the Kushites and Proto-Saharans into Eurasia from Middle Africa. This supports the proposition the R haplogroups originated in Africa, not Eurasia.
I agree with both, you and Swenet.
But the question I have, and maybe I overlooked this. You correlate V45 with "R-M207". But in the Fulvio Cruciani (2010) paper, he correlates V45 with M173.
Posted by Swenet (Member # 17303) on :
quote:Originally posted by Oshun:
quote:Originally posted by Swenet:
quote:Originally posted by the lioness,: Haplogroup R-V88 is thought to originated in thew Cameroon/Chad region
The 7ky old V88 Y chromosome in Iberia casts doubt on V88 originating in West/Central Africa. It can't be in West/Central Africa and Iberia at the same time so close to its estimated age. You're better off stating that most or all of the current variation of R-V88 comes from the Cameroon/Chad region.
I think you might have missed that extra information because I see that you quoted an older version of that post at the time that doesn't contain the caveats I put in there.
But weren't Iberians around this time Africans?
2005 Eva Fernández Domínguez
Nerja, Málaga (Spain)
NE-NM82.2 (Sample Name 2NE): Haplogroup J*: 17000-20000 NE-NAP (Sample Name 4NE): Haplogroup L1b: 2,260 BC NE-1829 (Sample Name 5NE): Haplogroup L1b: 5.875±80 A.P.
Toledo, Portugal K-13 (Sample Name: TO1): Haplogroup L3a: 9200-7800 A.P.
I wouldn't go as far as to say that contemporary Iberians as a whole were biologically Africans, just that they likely had substantially more African ancestry than contemporary mainland Neolithic and Bronze Age Europeans away from coastal areas in reach of Africans. La Brana also dates to this period and, aside from some what looks like African ancestry in his genome, he's definitely not African.
As I told Clyde, read the post I linked to for context of what I said. In that post I said that the Iberian R-V88 carrier might be a North African branch independent of the V88 that ended up going to West/Central Africa. So, I didn't bring up Iberian V88 to argue it originated there. I was trying to argue that it's unlikely that ALL North African V88 headed to West/Central Africa.
Remember that V88 got introduced to the ancestors of Chadic speakers at a time when Chadic might have still been a common language with, or at least in contact with, proto-Berber. In that case, some carriers could have stayed in North Africa, while some carriers went south. The southern branch could then have later migrated to North Africa again, explaining why R-V88 in Siwa Berbers and other North Africans looks at least partly like a West/Central African ancestry (as opposed to a North African one).
Posted by Clyde Winters (Member # 10129) on :
quote:Originally posted by Ish Gebor:
quote:Originally posted by Clyde Winters: Just because the oldest instance of a probable R1b-V88 in Iberia dates to 7,100ya (ID I0410, Els Trocs, Spain, Haak et al. 2015), does not make it older than the Chadian clade. Researchers have found that the TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010). Eurasians carry the M269 (R1b1b2) mutation. The subclades of R1b1b2 include Rh1b1b2g (U106) (TMRCA 8.3kya) and R1b1b2h (U152) (TMRCA 7.4kya). The most recent common ancestor for R1b1b2 is probably 8kya (Balaresque et al, 2010).
The fact that the earliest R1b aY-chromosome comes from Iberia should not be a surprise since West Africans frequently crossed into Eurasia via Iberia, instead of the Levant.
In Africa we find V45 (R-M207) and V88. Clearly, R-V88 in Chad is older than V88 in Iberia, Most importantly there is no archaeological evidence of a back migration of haplogroup R into Africa, but there is evidence of the migration of the Kushites and Proto-Saharans into Eurasia from Middle Africa. This supports the proposition the R haplogroups originated in Africa, not Eurasia.
I agree with both, you and Swenet.
But the question I have, and maybe I overlooked this. You correlate V45 with "R-M207". But in the Fulvio Cruciani (2010) paper, he correlates V45 with M173.
This is misdirection. If you don't keep up with the literature you want know what haplogroup is what. For example, mtDNA D, is the same as M1. They just change the name for Asian M1 to make it appear that M1 is isolated in Africa.
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by Clyde Winters:
quote:Originally posted by Ish Gebor:
quote:Originally posted by Clyde Winters: Just because the oldest instance of a probable R1b-V88 in Iberia dates to 7,100ya (ID I0410, Els Trocs, Spain, Haak et al. 2015), does not make it older than the Chadian clade. Researchers have found that the TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010). Eurasians carry the M269 (R1b1b2) mutation. The subclades of R1b1b2 include Rh1b1b2g (U106) (TMRCA 8.3kya) and R1b1b2h (U152) (TMRCA 7.4kya). The most recent common ancestor for R1b1b2 is probably 8kya (Balaresque et al, 2010).
The fact that the earliest R1b aY-chromosome comes from Iberia should not be a surprise since West Africans frequently crossed into Eurasia via Iberia, instead of the Levant.
In Africa we find V45 (R-M207) and V88. Clearly, R-V88 in Chad is older than V88 in Iberia, Most importantly there is no archaeological evidence of a back migration of haplogroup R into Africa, but there is evidence of the migration of the Kushites and Proto-Saharans into Eurasia from Middle Africa. This supports the proposition the R haplogroups originated in Africa, not Eurasia.
I agree with both, you and Swenet.
But the question I have, and maybe I overlooked this. You correlate V45 with "R-M207". But in the Fulvio Cruciani (2010) paper, he correlates V45 with M173.
This is misdirection. If you don't keep up with the literature you want know what haplogroup is what. For example, mtDNA D, is the same as M1. They just change the name for Asian M1 to make it appear that M1 is isolated in Africa.
In his earlier, 2003 paper he wrote:
quote: With the exception of a single Y chromosome from Morocco with the M269 mutation (haplotype 117b), all group IX African chromosomes are characterized by the presence of the M173 and M207 derived alleles and the absence of the downstream mutations (haplotype 117). Haplotype 117 was found only in Cameroon, where it accounts for 26% of the chromosomes (40% in northern Cameroon).
--Fulvio Cruciani
Am J Hum Genet. 2002 May; 70(5): 11971214. Published online 2002 Mar 21. doi: 10.1086/340257 PMCID: PMC447595
A Back Migration from Asia to Sub-Saharan Africa Is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes
It is as if people have been riding this horse ever since ( or for political reasons have been beating this death horse) and he, Fulvio, continued to move on.
But if correct you're saying that R-M207 equals M173, right?
Reading his paper again:
quote: These are: (1) haplotype 12b, carrying the M218 mutation but ancestral at M109; (2) haplotype 19b, carrying the derived alleles at M236 and M288 but ancestral at M146; (3) haplotype 41b, carrying the mutated allele at M98 but ancestral at M85; and (4) haplotype 117b, carrying the M269 derived allele on a M207/M173 background.
In his 2011 paper, Fulvio indeed correlates CT chromosomes with V45. So theoretically CT replaced M207.
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by Swenet:
quote:Originally posted by Oshun:
quote:Originally posted by Swenet:
quote:Originally posted by the lioness,: Haplogroup R-V88 is thought to originated in thew Cameroon/Chad region
The 7ky old V88 Y chromosome in Iberia casts doubt on V88 originating in West/Central Africa. It can't be in West/Central Africa and Iberia at the same time so close to its estimated age. You're better off stating that most or all of the current variation of R-V88 comes from the Cameroon/Chad region.
I think you might have missed that extra information because I see that you quoted an older version of that post at the time that doesn't contain the caveats I put in there.
But weren't Iberians around this time Africans?
2005 Eva Fernández Domínguez
Nerja, Málaga (Spain)
NE-NM82.2 (Sample Name 2NE): Haplogroup J*: 17000-20000 NE-NAP (Sample Name 4NE): Haplogroup L1b: 2,260 BC NE-1829 (Sample Name 5NE): Haplogroup L1b: 5.875±80 A.P.
Toledo, Portugal K-13 (Sample Name: TO1): Haplogroup L3a: 9200-7800 A.P.
I wouldn't go as far as to say that contemporary Iberians as a whole are biologically Africans, just that they likely had substantially more African ancestry than contemporary mainland Bronze Age Europeans away from coastal areas in reach of Africans.
As I told Clyde, read the post I linked to for context of what I said. In that post I said that the Iberian R-V88 carrier might be a North African branch independent of the V88 that ended up going to West/Central Africa. So, I didn't bring up Iberian V88 to argue it originated there. I was trying to argue that it's unlikely that ALL North African V88 headed to West/Central Africa.
Remember that V88 got introduced to the ancestors of Chadic speakers at a time when Chadic and Berber might have still been a common language with, or at least in contact with, proto-Berber. In that case, some carriers could have stayed in North Africa, while some carriers went south. The southern branch could then have later migrated to North Africa again, explaining why R-V88 in Siwa Berbers and other North Africans looks at least partly like a West/Central African ancestry (as opposed to a North African one).
Swenet, do you think some of these L* markers (posted by Oshun ) could relate to Southern Tuareg populations?
quote:The haplogroup R1b1a (R-V88) was found with a frequency of 8% in the village of Al Awaynat. Generally, haplogroup R and its subsets are spread in Eurasia as far as Siberia (Karafet et al., 2008; Chiaroni et al., 2009; Lancaster, 2010). Nevertheless, R1b1a has been observed at high frequencies in Northwest Africa (27% in the Egyptian Berbers), with peaks in the Chadic-speaking populations from Central Africa, ranging from 29 to 96% in Cameroon, and very rarely is found outside Africa (Cruciani et al., 2010a,b). This haplogroup has been proposed to represent the paternal genetic signature of the mid-Holocene migration of proto-Chadic Afro-asiatic speakers across the Central Sahara to Lake Chad (Ehret, 2002; Cruciani et al., 2010a); this suggests a link between Chadic speakers and other Afro-Asiatic speakers to the north of the Sahara.
In the eight-microsatellite Network analysis of R1b1a chromosomes from Northern and Central Africa (Fig. S2), the Libyan Tuareg R1b1a Y-chromosomes were found to belong to a branch characterized exclusively by haplotypes from Central Africa, more particularly from the Chad area (Cruciani et al., 2010a).
This may be likely explained by recent introduction through the slavery practices mentioned above.
Nonetheless, the hypothesis that the Libyan Tuareg R1b1a haplotypes may be relics of the migration of Pastoral proto-Chadic speakers, as hypothesized by the trans-Saharan hypothesis (Ehret, 2002; Cruciani et al., 2010a), cannot be ruled out
--Claudio Ottoni, et al. (2011)
Deep Into the Roots of the Libyan Tuareg: A Genetic Survey of Their Paternal Heritage
Posted by Ish Gebor (Member # 18264) on :
quote:Originally posted by Oshun:
quote:Originally posted by Swenet:
quote:Originally posted by the lioness,: Haplogroup R-V88 is thought to originated in thew Cameroon/Chad region
The 7ky old V88 Y chromosome in Iberia casts doubt on V88 originating in West/Central Africa. It can't be in West/Central Africa and Iberia at the same time so close to its estimated age. You're better off stating that most or all of the current variation of R-V88 comes from the Cameroon/Chad region.
I think you might have missed that extra information because I see that you quoted an older version of that post at the time that doesn't contain the caveats I put in there.
But weren't Iberians around this time Africans?
2005 Eva Fernández Domínguez
Nerja, Málaga (Spain)
NE-NM82.2 (Sample Name 2NE): Haplogroup J*: 17000-20000 NE-NAP (Sample Name 4NE): Haplogroup L1b: 2,260 BC NE-1829 (Sample Name 5NE): Haplogroup L1b: 5.875±80 A.P.
quote:"However, gene flow between sub-Saharan and northern African populations would also have been made possible earlier through the greening of the Sahara resulting from Early Holocene climatic improvement."
--Elika Podgorná et al.
Annals of Human Genetics Volume 77, Issue 6, pages 513523, November 2013
The Genetic Impact of the Lake Chad Basin Population in North Africa as Documented by Mitochondrial Diversity and Internal Variation of the L3e5 Haplogroup
quote:"The presence of almost 50% of sub-Saharan lineages L1b, L2 and L3 in Abauntz Chalcolithic deposits and Tres Montes, in Navarre, suggests the existence of an important gene flow from Africa to this geographic region. The low frequency of these lineages in the current Spanish population indicates that it has gene produced a replacement from the Chalcolithic period. The entry of African lineages could occur during the Paleolithic, during the Neolithic period, or during both periods. The phylogenetically related sequences present in the Chalcolithic deposit Iberian Peninsula and Neolithic and Chalcolithic samples of the Middle East points to Neolithic as most likely time of entry into the peninsula of these lineages."
--Fernández Domínguez, E. (2005)
Polimorfismos de DNA mitocondrial en poblaciones antiguas de la cuenca mediterránea.
Posted by Swenet (Member # 17303) on :
quote:Originally posted by Ish Gebor:
quote:Originally posted by Swenet:
quote:Originally posted by the lioness,: Haplogroup R-V88 is thought to originated in thew Cameroon/Chad region
The 7ky old V88 Y chromosome in Iberia casts doubt on V88 originating in West/Central Africa. It can't be in West/Central Africa and Iberia at the same time so close to its estimated age. You're better off stating that most or all of the current variation of R-V88 comes from the Cameroon/Chad region.
I think you might have missed that extra information because I see that you quoted an older version of that post at the time that doesn't contain the caveats I put in there.
Swenet,
What are the mtDNA L* markers correlating with these West / Central African populations?
Some of the more important ones with a West/Central African origin (except for L0):
quote:"Among the 16 expansion haplogroups, 5 lineages (L0a1a2, L2a1f, L3b1a1, L3e2a1b, and L3e3b) showed coalescence time less than 10 kya at least by three of five estimates mentioned in Methods, while the remaining 11 lineages (L0a1a, L1b1a3, L1b1a, L2a1a, L2a1c, L2a1, L3b1a, L3e1, L3e2a, L3e2b and L3d15) expanded before 10 kya (see Table S2). Specially, the expansion of lineage L3d15 took place before the LGM. From the ages estimated above, we found that most lineages shared among populations (11/15) expanded before 10 kya, i.e. the first occurrence of farming in the land of Africa. Thus, the result of lineage expansions showed that maternal African growth could be mainly attributed to pre-Neolithic expansion."
^ Thanks bro,
Posted by Doug M (Member # 7650) on :
I think that given the number of articles claiming that 'Eurasians' have significant mixture with Denisovans and Neanderthals, then if what they are saying about Eurasian mixture in Africa is true, then they should also see the Neanderthal and Denisovan DNA in these African populations.
But they don't......
Posted by Djehuti (Member # 6698) on :
^ That also depends on what genes one may even consider to be of another hominid species. Recall the claims of neanderthal ancestry in Maasai!!
Posted by Elmaestro (Member # 22566) on :
quote:Originally posted by Doug M: I think that given the number of articles claiming that 'Eurasians' have significant mixture with Denisovans and Neanderthals, then if what they are saying about Eurasian mixture in Africa is true, then they should also see the Neanderthal and Denisovan DNA in these African populations.
Slowly but surely Ne/Den admixture is popping up in every corner of the dark continent... Which is my concern, I know for a fact that more and more and more SSA populations would display some form of admixture previously ruled out, eventually. It will come down to how we intemperate the data individually.
Posted by Swenet (Member # 17303) on :
Doug, see fig 5.
Anyone able to see and point out what needs to be said about fig 5?
Posted by Ish Gebor (Member # 18264) on :
quote:
Haplogroup A1b. The P114 mutation, which defines hap- logroup A1b according to Karafet et al. [14], had been detected in central-western Africa at very low frequencies (in total, three chromosomes from Cameroon) [16,19].
[...]
Out of Africa haplogroups. All Y-clades that are not exclusively African belong to the macro-haplogroup CT, which is defined by mutations M168, M294 and P9.1 [14,31] and is subdivided into two major clades, DE and CF [1,14]. In a recent study [16], sequencing of two chromosomes belonging to haplogroups C and R, led to the identification of 25 new mutations, eleven of which were in the C-chromosome and seven in the R-chromosome. Here, the seven mutations which were found to be shared by chromosomes of haplogroups C and R [16], were also found to be present in one DE sample (sample 33 in Table S1), and positioned at the root of macro-haplogroup CT (Figure 1 and Figure S1). Six haplogroup C chromosomes (samples 3439 in Table S1) were analyzed for the eleven haplogroup C- specific mutations [16] and for SNPs defining branches C1 to C6 in the tree by Karafet et al. [14] (Figure S1). Through this analysis we identified a chromosome from southern Europe as a new deep branch within haplogroup C (C-V20 or C7, Figure S1). Previously, only a few examples of C chromosomes (only defined by the marker RPS4Y711) had been found in southern Europe [32,33]. To improve our knowledge regarding the distribution of haplogroup C in Europe, we surveyed 1965 European subjects for the mutation RPS4Y711 and identified one additional haplogroup C chromosome from southern Europe, which has also been classified as C7 (data not shown). Further studies are needed to establish whether C7 chromosomes are the relics of an ancient European gene pool or the signal of a recent geographical spread from Asia. Two mutations, V248 and V87, which had never been previously described, were found to be specific to haplogroups C2 and C3, respectively (Figure S1). Three of the seven R-specific mutations (V45, V69 and V88) were previously mapped within haplogroup R [34], whereas the remaining four mutations have been here positioned at the root of haplogroups F (V186 and V205), K (V104) and P (V231) (Figure S1) through the analysis of 12 haplogroup F samples (samples 4051, in Table S1).
[...]
Supporting Information
Figure S1 Structure of the macro-haplogroup CT. For details on mutations see legend to Figure 1. Dashed lines indicate putative branchings (no positive control available). The position of V248 (haplogroup C2) and V87 (haplogroup C3) compared to mutations that define internal branches was not determined. Note that mutations V45, V69 and V88 have been previously mapped (Cruciani et al. 2010; Eur J Hum Genet 18:800807). (TIF)
--Fulvio Cruciani et al.
Molecular Dissection of the Basal Clades in the Human Y Chromosome Phylogenetic Tree
quote:
To test the robustness of the backbone and the root of current Y chromosome phylogeny, we searched for SNPs that might be informative in this respect. To this aim, a resequencing analysis of a 205.9 kb MSY portion (183.5 kb in the X-degenerate and 22.4 kb in the X-transposed region) was performed for each of seven chromosomes that are representative of clade A (four chromosomes belonging to haplogroups A1a, A1b, A2, and A3), clade B, and clade CT (two chromosomes belonging to haplogroups C and R) (Table S1 available online).
The phylogenetic relationships we observed among chromosomes belonging to haplogroups B, C, and R are reminiscent of those reported in the tree by Karafet et al.13 These chromosomes belong to a clade (haplogroup BT) in which chromosomes C and R share a common ancestor (Figure 2).
--Fulvio Cruciani
A Revised Root for the Human Y Chromosomal Phylogenetic Tree: The Origin of Patrilineal Diversity in Africa
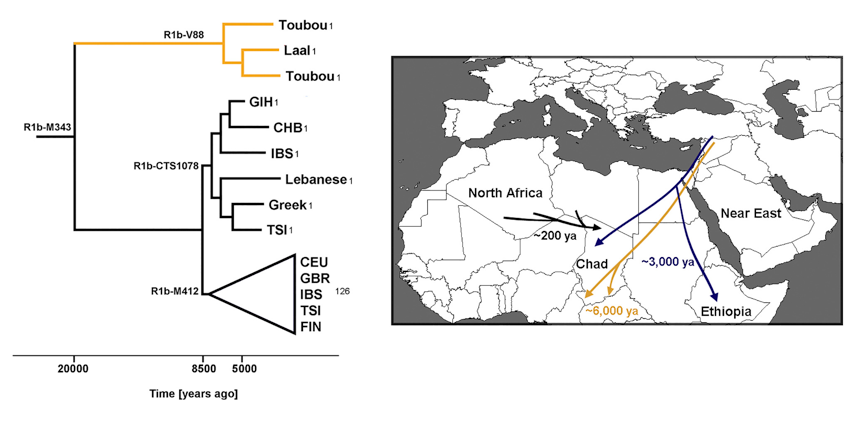



![[Smile]](smile.gif)
![[Confused]](confused.gif)
![[Wink]](wink.gif)