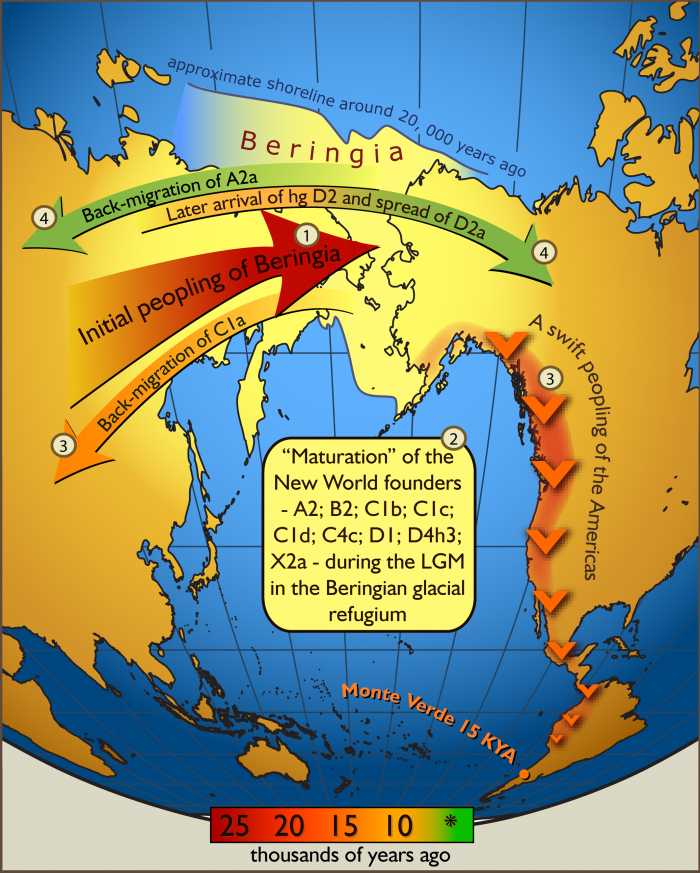
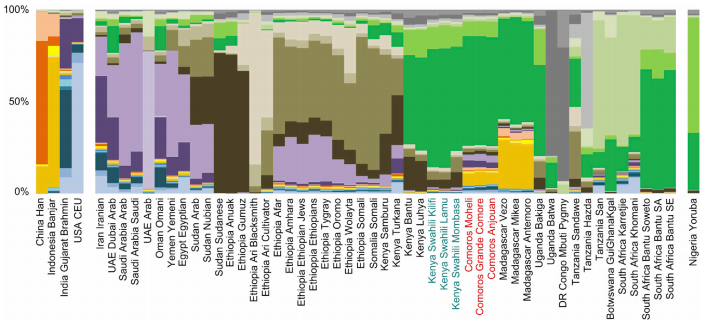
code:
# Primary Population (source) Secondary Population (source) Distance
1 66.1% Hadza + 33.9% Levant_N @ 5.39
2 64.3% Hadza + 35.7% Levant_BA @ 5.62
3 74.9% Masai + 25.1% Levant_N @ 5.91
4 61.8% Mota + 38.2% Levant_BA @ 6.78
5 63.6% Mota + 36.4% Levant_N @ 6.89
6 73.6% Masai + 26.4% Levant_BA @ 7.09
7 51.5% Gambian + 48.5% Levant_BA @ 7.95
8 50.1% Levant_BA+ 49.9% Esan @ 8.16
9 50.1% Levant_BA+ 49.9% Yoruba @ 8.16
10 60.1% Masai + 39.9% Moroccan @ 8.21
11 66.2% Masai + 33.8% Libyan @ 8.73
12 61.2% Masai + 38.8% Saharawi @ 8.74
13 50.2% Hadza + 49.8% Saharawi @ 8.75
14 72.8% Masai + 27.2% Jew_Libyan @ 9.19
15 67.9% Masai + 32.1% Egyptian @ 9.3
16 55.6% Hadza + 44.4% Libyan @ 9.3
17 77.9% Masai + 22.1% Anatolia_ChL @ 9.31
18 73.3% Masai + 26.7% Jew_Tunisian @ 9.37
19 52.8% Saharawi+ 47.2% Mota @ 9.47
20 63.5% Masai + 36.5% Algerian @ 9.48

![[Smile]](smile.gif)



![[Big Grin]](biggrin.gif)
![[Frown]](frown.gif)
![[Embarrassed]](redface.gif)
![[Confused]](confused.gif)
![[Wink]](wink.gif)

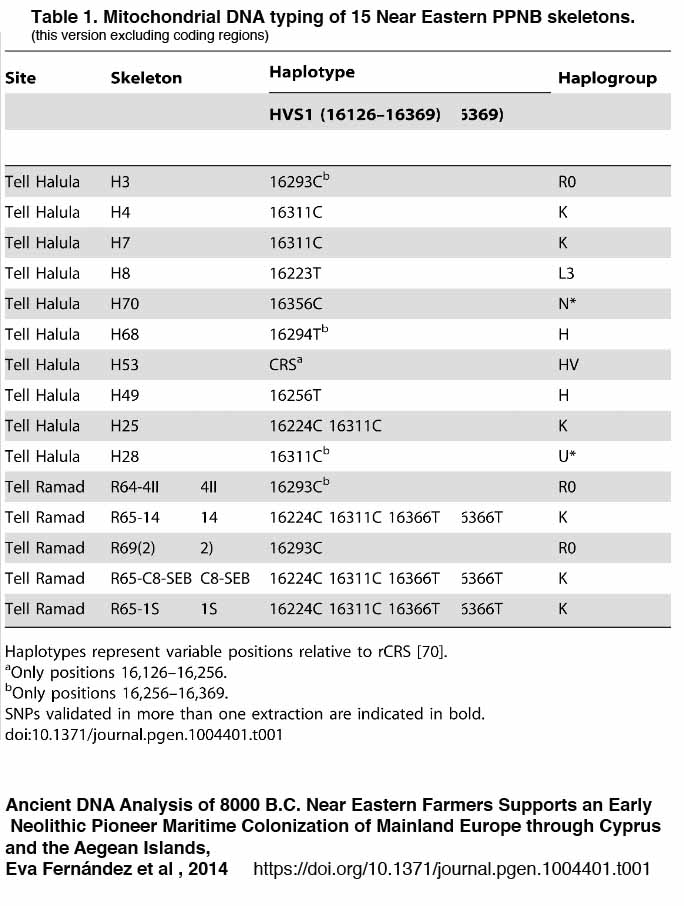
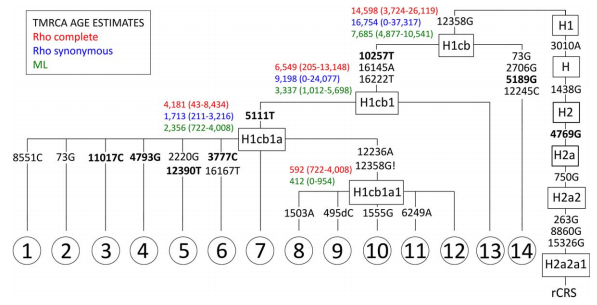
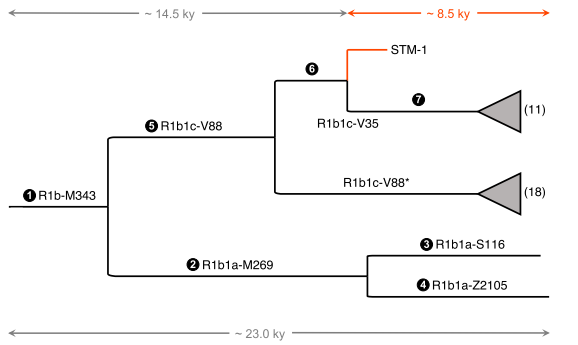
![[Roll Eyes]](rolleyes.gif) In fact this Soqotri N* is I5a according to Fernandes et al 2012, I5a2a on PhyloTree. The Algerian N from the third source is some other random subclade unrelated to the Yemeni one.
In fact this Soqotri N* is I5a according to Fernandes et al 2012, I5a2a on PhyloTree. The Algerian N from the third source is some other random subclade unrelated to the Yemeni one.