Abstract The origin and evolution of modern humans in Africa has reached a multidisciplinary consensus but the age and regions where it originated and evolved are current topics of discussion. In this study I put forward an integrative model guided by the phylogeny and phylogeography of mitochondrial DNA (and Y-chromosome) haplogroups. I propose an early origin of modern humans in northwest Africa in a temporal window of 257-345 thousand years ago. A first population split in central Africa around 175-288 thousand years ago. A subsequent northward spread with additional population subdivisions during a long statistical interval that culminated in a first successful out of Africa migration around 130 thousand years ago. A population constriction in southwest Asia motivated an early return to Africa between 79 and 107 thousand years ago. This ample Eurasian-ebb to Africa, detected by mitochondrial haplogroup L3 and Y-chromosome haplogroup E preceded other later and geographically more limited Eurasian backflows. The archaeological and fossil finds that could be coetaneous to this molecular journey have been integrated into this interdisciplinary model.
__________________________
The northern African origin of the ancestor of modern humans and Neanderthals
Based on the topologies obtained from non-recombinant uniparental markers (Green et al. 2008; Mendez et al. 2016; Meyer et al. 2012, 2014; Petr et al. 2020)[35,36,4749], I have proposed recently that modern humans and Neanderthals were sister clades [50], and that the topologies obtained using autosomal markers [37,48,51], which consider Homo sapiens as an outgroup of the sister pair Neanderthal-Denisovan were due to secondary introgression. Furthermore, I also posit that the ancestor of modern humans and Neanderthals originated in northern Africa, and that pre-Neanderthal groups crossed to Europe whereas the ancestors of modern humans remained in northern Africa, so that both groups evolved in allopatry [50].
The northwest African origin of early anatomically modern humans
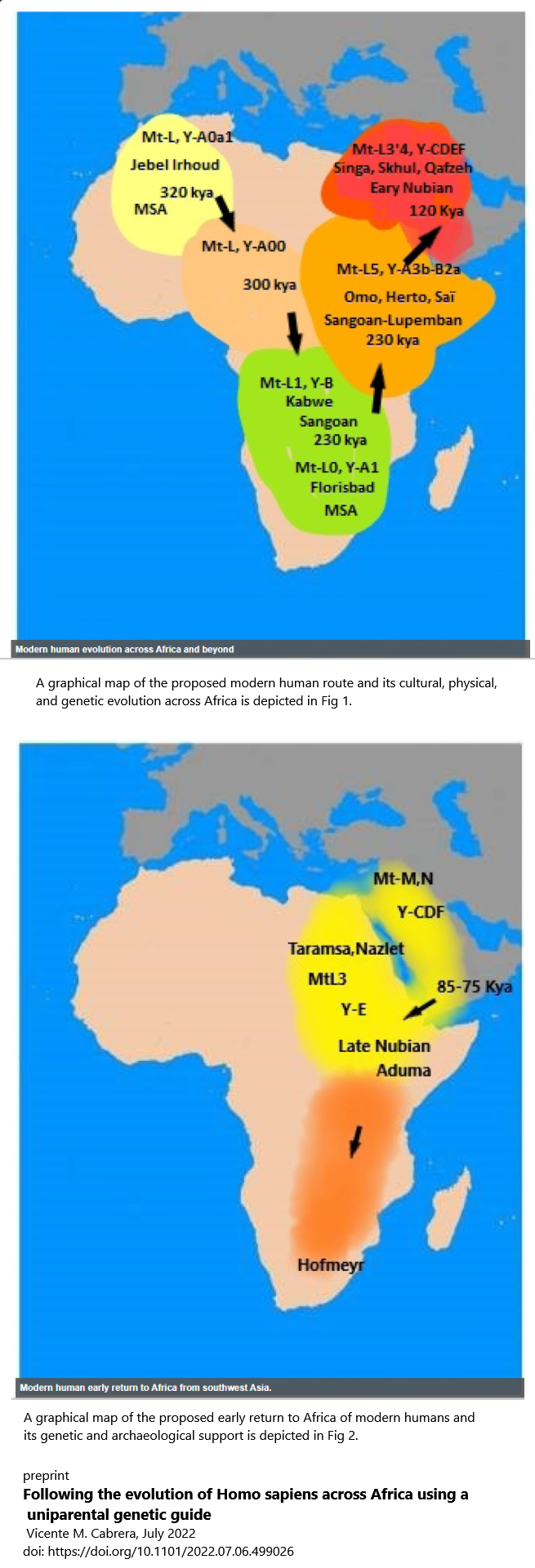
Human fossil remains and Middle Stone Age (MSA) archaeological artefacts from Jebel Irhoud, Morocco, dated at 315 ± 34 thousand years ago (Kya) have situated the earliest phase of modern human evolution in northwest Africa [4,5]. Applying a variable evolutionary rate dependent of temporal fluctuations in population size to the mtDNA genome, a coalescent age for the most recent common ancestor of all extant mtDNA lineages was estimated around 300 kya [14,21] which has been replicated in this study (Table 1).
This mtDNA coalescence matches the archaeological and fossil estimates in Morocco but there is a lack of specific mtDNA lineages in this area to directly support a northwest African origin. However we have indirect evidence of the existence of an old genetic component in the Maghreb. Thus, Late Pleistocene northern African remains derived one-third of their genomic ancestry from a complex sub-Saharan African gene pool [52]. Curiously, this component was not detected in subsequent Neolithic periods [53]. On the other hand, it is interesting to point out that, although most of the Y-chromosome lineages in Morocco (J-M267; E-M81) are of recent implantation [54], one of the most ancient lineages of the Y-Chromosome, A0a1 (xP114) has been detected in Moroccan Berbers [55]. Accepting the northwest African origin hypothesis implies that other contemporaneous hominin lineages as the Broken Hill (Zambia) skull dated to 299 ± 25 kya [56], or the Kenyan Guomde calvarium dated to around 270 kya [57] possibly did not directly contribute to the origin of our species.
It has been proposed that after the radiation of L3 in eastern Africa, the ancestors of M and N crossed the Bab al Mandab strait about 60 70 Kya (the previously calculated coalescent age for L3) and, following a southern coastal route, they spread all over the world [100,104106]. As an alternative hypothesis, we have proposed that the clade L346 already extended its geographic range to southwestern Asia and that the splits of the L6 and L34 branches (Table 2) occurred at the outside margins of Africa, being the Y-chromosome counterpart of this early spread the haplogroup CT-M168 that includes the Eurasian haplogroups C, D and F and the African haplogroup E[23]. The ample statistical range of these mtDNA coalescent ages (Table 2) includes important archaeological finds in the region as the presence of early modern human populations in the Levant at Misliya Cave from 177 to 194 kya[8], and at Qafzeh[107,108] and Skhul[109,110] caves from 90 to 130 kya.
An earlier return to Africa
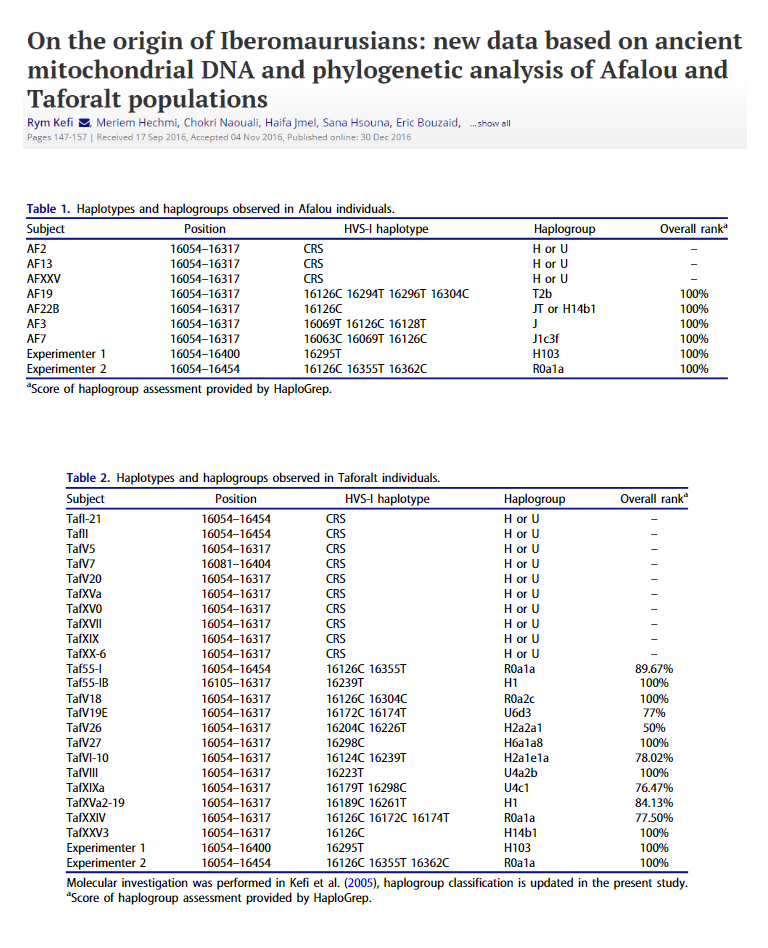
After a period of maturation and stasis in southwestern Asia, mtDNA haplogroup L3 split in the region and while the ancestors of the L3 African subclades returned to Africa, the ancestors of the Eurasian branches M and N began their exodus eastwards[23]. According to the new coalescent ages for the L3 subclades (Table 3), the first radiations in eastern Africa took place around 75 kya, at the beginning of the arid MIS 4 period. It was at this stage when an early modern human displacement by the Neanderthals in the Levant was attested [40]. The Y-chromosome counterpart of this mtDNA back flow to Africa was haplogroup E [127]. The detection, in the extant population of Saudi Arabia, of the basal African Y-chromosome lineage E-M96*[128] is in support of this back flow. Furthermore, whole genome sequence analyses also favor models involving possible African returns 70-60 kya[129,130]. Interestingly, a similar model, involving back flow to Africa, has been proposed to explain the complex mtDNA phylogeography of hamadryas baboons lineages present in Africa and Arabia [131].
The evidence gathered from the fossil and archaeological records for the proposed return to Africa has been only occasionally mentioned but, without generalized acceptance. Thus, it has been suggested that the Early Nubian Complex, developed at the end of MIS 6 beginning of MIS5 (145 125 Kya) in northeast Africa, extended to Arabia where the Late Nubian Complex occurred and from there was reintroduced into Africa during MIS5a (85-75 kya)
The first back to Africa of modern humans
The first out of Africa and back again for modern humans was proposed based on a nested cladistic analysis of the Y-chromosome variation (Hammer [159], and was supported by applying a most parsimonious criterion at an unbiased Y-chromosome tree [127]. Searching for a female counterpart, it was suggested that mtDNA haplogroup L3 also signals an early return to Africa [23] and, recently, this backflow to Africa has also been detected by whole genomic data [129]. The relatively closer morphological affinities of some African fossils with coetaneous Eurasian remains rather than with current African groups that have never abandoned the African continent [150], could also be taken in favor of this return to Africa. However, the archaeological support is much weaker because, although the temporal margins of the appearance and development of the Early and Late Nubian technological complexes are into the range proposed by the genetic markers, a clear geographical and temporal separation between these two lithic variants have not been yet determined. Therefore, it remains to deepen into this possibility suggested only by a few [132].
keep in mind discussion of L3'4'6 in the article look at the timeline above, oldest at top L3'4'6 is older than L3'4 and both of them are older than L3 L5 is older than L2 and L3, etc
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
posted
if you look at the bolded in the abstract and the last paragraph above which is also the last paragraph of the article one o the main themes of this article is back to Africa" migration and they mainly discuss that by analysis of mitochondrial L3 and this they pair with Y DNA A as we can see on the lower map, this proposed back flow is very old 85-75 kya and coming in around Ethiopia, the arrow shows. This is not the same relatively much more recent back flows discussed in other articles such as 2012's :
Genomic Ancestry of North Africans Supports Back-to-Africa Migrations Brenna M. Henn
Abstract
We identify a gradient of likely autochthonous Maghrebi ancestry that increases from east to west across northern Africa; this ancestry is likely derived from back-to-Africa gene flow more than 12,000 years ago [ya], prior to the Holocene.
quote:it should be mentioned the presence of a Sangoan of northeastern Africa technology included over a northwestern Africa Levallois Mousterian substratum at Wadi Lazalim in southern Tunisia [97]. Afterward, molecular markers signal a clear northward geographic progression signaled by L5 and L346 mtDNA clades at the eastern African region and, less evidently, by the L2 clade at the central region. In northeastern Africa it seems that the sub-Saharan Sangoan/Lupemban was replaced by the Early Nubian technology [132]. It is also probable that in northern Africa it was the Aterian which evolved from previous sub-Saharan lithic industries [151]. Nevertheless, the out of African migrants carrying maternal clade L346 and paternal clade CT-M168 only could brought an Early Levantine Mousterian industry to the Levant and a possible related Lupemban technology to southern Arabia [117] and, afterwards, an Early Nubian technology that spread and differentiated across the whole peninsula [152]. These early demic spreads out of Africa into Eurasia, coinciding with humid periods as the end of MIS7 (around 190 ka) and MIS5e (around 130 kya), could satisfactorily explain the detection of anatomically modern human teeth in southern China dated to 120-80 Kya[9], the presence of an early modern human tooth in Sumatra at 73-63 kya[153], the archaeological evidence of a possible human arrival to northern Australia around 65 kya [154], or the genomic evidence of an ancient split between Africans and Papuans around 120 kya[10].
Interestingly while several Taforalt individuals were of haplogroup E of the about 30 individual both the Afalou in Algeria and the Moroccan Taforalt, none on this chart were of mtDNA L
In the OP that top map in yellow they have marked
quote: Mt L, Y A0a1
Jebel Irhoud
320 Kya
So this uniparental aspect is not L with E it's L with A they are speculating here since there was no analyzable DNA found at Jebel Irhoud
they also make reference to
quote: " On the other hand, it is interesting to point out that, although most of the Y-chromosome lineages in Morocco (J-M267; E-M81) are of recent implantation [54], one of the most ancient lineages of the Y-Chromosome, A0a1 (xP114) has been detected in Moroccan Berbers [55]. " there are other Y-chromosome basal lineages, as A0a1a observed in Cameroonian Bakola directly related to the A0a1 (xP114) present in Berbers from Algeria, or the primitive A1a clade observed in Fulbe and Tuareg from Niger and also found in Moroccan Berbers, that consistently points to an early migratory input from northwest to western Africa[64]
_________________
[55]55.↵Cruciani F, Trombetta B, Massaia A, Destro-Bisol G, Sellitto D, Scozzari R. A revised root for the human Y chromosomal phylogenetic tree: the origin of patrilineal diversity in Africa. The American Journal of Human Genetics. Elsevier; 2011;88(6):8148.Google Scholar
Four subjects (two Berbers from northwest Africa, one Tuareg and one Fulbe from Niger) were confirmed as belonging to clade A1a.
Tukuler
multidisciplinary Black Scholar
Member # 19944
posted
quote:
M. van de Loosdrecht, A. Bouzouggar, L. Humphrey, C. Posth, N. Barton, A. Aximu-Petri, B. Nickel, S. Nagel, E.H. Talbi, M.A. El Hajraoui, et al. Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations Science, 360 (2018), pp. 548-552 https://www.science.org/cms/asset/631b5cb2-9bd2-49ee-9544-4e3b9313eb23/pap.pdf
The relationships of the Iberomaurusian culture with the preceding MSA, including the local backed bladelet technologies in Northeast Africa, and the Epigravettian in southern Europe have been questioned (13). The genetic profile of Taforalt suggests substantial Natufian-related and sub-Saharan African-related ancestries (63.5% and 36.5%, respectively), but not additional ancestry from Epigravettian or other Upper Paleolithic European populations. Therefore, we provide genomic evidence for a Late Pleistocene connection between North Africa and the Near East, predating the Neolithic transition by at least four millennia, while rejecting a potential Epigravettian gene flow from southern Europe into northern Africa within the resolution of our data. Archaeogenetic studies on additional Iberomaurusian sites will be critical to evaluate the representativeness of Taforalt for the Iberomaurusian gene pool. We speculate that the Natufian related ancestral population may have been widespread across North Africa and the Near East, associated with microlithic backed bladelet technologies that started to spread out in this area by at least 25,000 yBP ((10) and references therein). However, given the absence of ancient genome data from a similar time frame for this broader area, the epicenter of expansion, if there was any, for this ancestral population remains unknown.
Supplement https://www.science.org/doi/10.1126/science.aar8380 S5. An overview of the genetic affinity of Taforalt with worldwide populations based on PCA and ADMIXTURE Marieke van de Loosdrecht, Choongwon Jeong
S6. Genetic affinity of Taforalt with early Holocene Levantines and present-day sub-Saharan Africans Marieke van de Loosdrecht*, Choongwon Jeong*
[ You'll have to goto the PDF for this one. Too technical for overview snippetting. ]
From FIG. S22 Six Taforalt mitogenomes fall on the U6a branch and one on the M1b branch. M1b TAF014.A0302 (Marocco 14.5ka) U6a TAF009.A0301 (Marocco 14.5ka) U6a TAF015.A0401 (Marocco 14.5ka) U6a TAF013.A0302 (Marocco 14.5ka) U6a TAF010.A0302 (Marocco 14.5ka) U6a TAF011.A0402 (Marocco 14.5ka) U6a TAF012.A0301 (Marocco 14.5ka)
S12. Y-chromosome haplogroup assignment Choongwon Jeong, Marieke van de Loosdrecht
In all six males, we observe haplogroup E1b1b, more specifically E1b1b1a1 (M-78) in five of six (Table S16). This haplogroup is most frequent in present-day North and Northeast African populations, such as Oromo, Somali and Moroccan Arabs (18). A previous study reported that Natufians and Neolithic Levant individuals had E1b1b haplogroups, although they tended to belong to E1b1b1b (16).
From Table S16. Y-haplogroup assignment for six Taforalt males. All individuals could be assigned to haplogroup E1b1b, and five of them more 1273 specifically to E1b1b1a1 (M-78). TAF009 (E1b1b1a1b1) TAF010 (E1b1b1a1) TAF011 (E1b1b1a1) TAF013 (E1b1b1a1) TAF014 (E1b1b1a1) TAF015 (E1b1b)
_______________________________
Population history of North Africa based on modern and ancient genomes Marcel Lucas-Sánchez et al, 2021
posted
I think he is partly correct about the AMH origin in North West Africa. Though i would argue its a bit more complex than even that. Something i wrote last year.
quote: How about this: Anatomically Modern Human or Homo Sapiens is a composite species consisting of a North West African patrilineage in A00 and an East African matrilineage in L0.
Thus Jebel Irhoud in Morocco at 300-150 kya and Omo Kibbish in Ethiopia at 200kya - If we had their DNA would be totally different. These different elements, *actually their regional ancestors* would come together to create the stabilized mixture that would later be known as "Homo Sapiens".
A00 has an upper bound date of 500kya. L0 upper bond is half that at 250kya. They were not the only "Adam" and "Eve" but these are the ones that persisted and they dont come from the same place.
Another point of evidence: East Africa particularly Ethiopia has TONS of maternal diversity. When you look at haplogroup L, most of the daughter lineages are East African, There are a handful of East African specific L3's like L3i, L3x, L3h, L3a, L3f, and L3m and L3n. PLUS the East has L4, L5, L6, L7 - Everything maternally at an early date was popping.
The Earliest non East African maternal lineage is L1 at 100-140kya, 100kya AFTER things been popping in the East.
West Africa has tons of PATERNAL diversity: Oldest A, Oldest B, E1a, E2, DE*, E, D0, Its almost the opposite of what you see in the East when you look at EARLY paternal diversity.
DE* was found in WEST Africa, D0 was found in WEST Africa. A00, A0, A1a, Rare B1 and B2 in the Sahel other rare B in West Africa.
EDIT - Jsut finished reading it. This publications, like its other publications that made similar arguments fall flat in the face of the Genetic evidence. IF this theory was true we would see the mtdna diversity that we see in East Africa actually in West Asia. There are not West Asian specific clades of L3 other than M and N and those L3 lineages are not OLDER than African L3 lineages like L3h. There is D0 in Africa. And DE* IN Africa - All the way to the WEST of Africa far away from West Asian These lineages of DE and D0 in West Africa already precede the date they envision for Back Migration.
Not surprised there is only ONE name on this study. He does a lot of mental gymnastics to argue these lineages originate in Asia....but they didn't survive there...and all the evidence we have of their existence is actually in Africa....but they not African.
Was it Diop that stated "Thus we reach the absurd conclusion that Blacks are basically Whites"
posted
I'm noticing that Vicente M. Cabrera the author of the article in the OP had a very similar article in 2018 (below) that we commented on in 2018, This new one in 2022 is virtually repeat but he was more explicit in the 2018 version
It is likely that the divergence of the Y-chromosome D and E haplogroups occurred up to the Himalayas and in or westward of the Tibet which coincides with the hypothetical bifurcation center proposed for the mtDNA L3 macrohaplogroup. (last sentence)
quote:Originally posted by beyoku:
Was it Diop that stated "Thus we reach the absurd conclusion that Blacks are basically Whites"
Actually the Asiatic Black man
(unless you're Twa)
Thus most AAs (not including Chris Rock) are actually Asian Americans according to Cabrera/NOI
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
Table 2) was detected in Mali. Although it shares the HVI 222 transition with other North African sequences belonging to haplogroup H (Rando et al. 1998), the RFLP analysis placed it in the basal HV cluster. Surprisingly for a western Africa country, around 42% of the sub-Saharan African sequences were L3 lineages. The predominant haplogroups belonged to L3b (17%), L3e (13%) and L3d (8%), but with different distribution within ethnolinguistc groups. Whereas the Bambara have higher frequencies of L3b (21%) and L3e (10%), L3d (10%) is similar in both samples...
~González AM,Mitochondrial DNA variation in Mauritania and Mali and their genetic relationship to other Western Africa populations.
quote:Within the human mitochondrial DNA (mtDNA) tree, haplogroup L3 encompasses not only many sub-Saharan Africans but also all ancient non-African lineages, and its age therefore provides an upper bound for the dispersal out of Africa. An analysis of 369 complete African L3 sequences places this maximum at ∼70 ka, virtually ruling out a successful exit before 74 ka, the date of the Toba volcanic supereruption in Sumatra.
[ ]
The L3 mtDNA pool within Africa suggests a migration from Eastern Africa to Central Africa ∼60 to 35 ka and major migrations in the immediate postglacial again linked to climate. The largest population size increase seen in the L3 data is 34 ka in Central Africa, corresponding to Bantu expansions, leading diverse L3 lineages to spread into Eastern and Southern Africa in the last 32 ka.
~Pedro Soares et al.,The Expansion of mtDNA Haplogroup L3 within and out of Africa Molecular Biology and Evolution, Volume 29, Issue 3, 1 March 2012, Pages 915927
quote:Originally posted by Ish Geber: [QB] By the logic implemented above,
quote:The Bambara & Madinka 40% L3 Lineage
Table 2) was detected in Mali. Although it shares the HVI 222 transition with other North African sequences belonging to haplogroup H (Rando et al. 1998), the RFLP analysis placed it in the basal HV cluster. Surprisingly for a western Africa country, around 42% of the sub-Saharan African sequences were L3 lineages. The predominant haplogroups belonged to L3b (17%), L3e (13%) and L3d (8%), but with different distribution within ethnolinguistc groups. Whereas the Bambara have higher frequencies of L3b (21%) and L3e (10%), L3d (10%) is similar in both samples...
~González AM,Mitochondrial DNA variation in Mauritania and Mali and their genetic relationship to other Western Africa populations.
^^^ Interestingly, the second coauthor for this article was also Vincent Cabrera (Universidad de La Laguna, Tenerife, Canary Islands)
quote: Mitochondrial DNA variation in Mauritania and Mali and their genetic relationship to other Western Africa populations 2006
A M González 1, V M Cabrera, J M Larruga, A Tounkara, G Noumsi, B N Thomas, J M Moulds Affiliations expand PMID: 16907709 DOI: 10.1111/j.1469-1809.2006.00259.x
Abstract Mitochondrial DNA (mtDNA) variation was analyzed in Mauritania and Mali, and compared to other West African samples covering the considerable geographic, ethnic and linguistic diversity of this region. The Mauritanian mtDNA profile shows that 55% of their lineages have a west Eurasian provenance, with the U6 cluster (17%) being the best represented. Only 6% of the sub-Saharan sequences belong to the L3A haplogroup a frequency similar to other Berber speaking groups but significantly different to the Arabic speaking North Africans. The historic Arab slave trade may be the main cause of this difference. Only one HV west Eurasian lineage has been detected in Mali but 40% of the sub-Saharan sequences belong to cluster L3A. The presence of L0a representatives demonstrates gene flow from eastern regions. Although both groups speak related dialects of the Mande branch, significant genetic differences exist between the Bambara and Malinke groups. The West African genetic variation is well structured by geography and language, but more detailed ethnolinguistic clustering suggest that geography is the main factor responsible for this differentiation.
the Bambara from Mali do not cluster with the other Mande speakers, including the Malinke who are also from Mali. The high frequency of L3b and presence of L0a lineages pulls the Bambara towards the eastern Hausa and Bassa.
As Bambara is also the dominant language in Mali these results suggest a cultural homogenizing effect on a genetically diverse background.
The area occupied by Cameroon is not always considered as part of the geographic region known as West Africa.
However, favouring a geographic continuum, the L3e haplogroup links Mali and Niger-Nigeria to Cameroon. Only L3e4 which is more abundant in the western countries is an exception. These results are in agreement with the Central African origin proposed for this haplogroup
wikipedia:
L3e Spread from East Africa in the upper paleolithic to West-Central Africa. It is the most common L3 sub-clade in Bantu-speaking populations.
[26] L3e is suggested to be associated with a Central African origin and is also the most common L3 subclade amongst African Americans, Afro-Brazilians and Caribbeans[27] ________________________
Haplogroup L3e is widespread in Africa but practically absent in Eurasia (except in neighbouring areas due to genetic exchange). Its origin is situated in Central or Eastern Africa about 46 Kya (Bandelt 2001)[83] and one of its most frequent lineages in West-Central Africa is L3e2b (7%) [84].
this is from the OP article Note the age of each clade in descending order but the names not corresponding These clade ages are pertinent to first map at top
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |






 UBBFriend: Email this page to someone!
UBBFriend: Email this page to someone!













 Printer-friendly view of this topic
Printer-friendly view of this topic