In the Table I cite the source and the frequency. Only a fool and moron would accept your statements as valid given the fact you have repeatedly shown that the figures are correct.
Stop trying to steal the history of Black people.
You can whirl , you can spin, you can emit lots of spam BUT there is no R-M269 in Africa, R-V88 (R1b1) is only present in a limited area in Africa, there is no R-M173 (R1) in Africa. There is nothing to refute. Something has to proven before it can be refuted. It is up to the proposer to produce the evidence for a proposal. You have only produced misquotes. Please quote word-for-word refereed papers that say "M-269 is widespread in Africa" "M-173 is widespread in Africa, "R=P25*(R1b1) is widespread in Africa.
quote: The Y chromosome Alu polymorphism (YAP, also called M1) defines the deep-rooted haplogroup D/E of the global Y-chromosome phylogeny [1]. This D/E haplogroup is further branched into three sub-haplogroups DE*, D and E (Figure 1). The distribution of the D/E haplogroup is highly regional, and the three subgroups are geographically restricted to certain areas, therefore informative in tracing human prehistory (Table 1). The sub-haplogroup DE*, presumably the most ancient lineage of the D/E haplogroup was only found in Africans from Nigeria [2], supporting the "Out of Africa" hypothesis about modern human origin. The sub-haplogroup E (E-M40), defined by M40/SRY4064 and M96, was also suggested originated in Africa [3-6], and later dispersed to Middle East and Europe about 20,000 years ago [3,4]. Interestingly, the sub-haplogroup D defined by M174 (D-M174) is East Asian specific with abundant appearance in Tibetan and Japanese (3040%), but rare in most of other East Asian populations and populations from regions bordering East Asia (Central Asia, North Asia and Middle East) (usually less than 5%) [5-7]. Under D-M174, Japanese belongs to a separate sub-lineage defined by several mutations (e.g. M55, M57 and M64 etc.), which is different from those in Tibetans implicating relatively deep divergence between them [1]. The fragmented distribution of D-M174 in East Asia seems not consistent with the pattern of other East Asian specific lineages, i.e. O3-M122, O1-M119 and O2-M95 under haplogroup O [8,9].
quote: Further refinement awaits the finding of new markers especially within paragroup E3a*-M2. The microsatellite profile of the DE* individual is one mutational step away from the allelic state described for Nigerians (DYS390*21, DYS388 not tested; [37], therefore suggesting a common ancestry but not elucidating the phylogenetics.
Haplogroup DE* in Guinea-Bissau:
Y-chromosomal diversity in the population of Guinea-Bissau: a multiethnic perspective
quote: There has been considerable debate on the geographic origin of the human Y chromosome Alu polymor- phism (YAP). Here we report a new, very rare deep-rooting haplogroup within the YAP clade, together with data on other deep-rooting YAP clades. The new haplogroup, found so far in only five Nigerians, is the least-derived YAP haplogroup according to currently known binary markers. However, because the interior branching order of the Y chromosome genealogical tree remains unknown, it is impossible to impute the origin of the YAP clade with certainty. We discuss the problems presented by rare deep-rooting lineages for Y chromosome phylogeography.
Haplogroup DE* in Nigerians:
Rare Deep-Rooting Y Chromosome Lineages in Humans: Lessons for Phylogeography
Y-DNA haplogroup R-M207 is believed to have arisen approximately 27,000 years ago in Asia. The two currently defined subclades are R1 and R2.
quote:Out of Africa haplogroups.
All Y-clades that are not exclusively African belong to the macro-haplogroup CT, which is defined by mutations M168, M294 and P9.1 [14,31] and is subdivided into two major clades, DE and CF [1,14].
In a recent study [16], sequencing of two chromosomes belonging to haplogroups C and R, led to the identification of 25 new mutations, eleven of which were in the C-chromosome and seven in the R-chromosome.
Here, the seven mutations which were found to be shared by chromosomes of haplogroups C and R [16], were also found to be present in one DE sample (sample 33 in Table S1), and positioned at the root of macro-haplogroup CT (Figure 1 and Figure S1)
Structure of the macro-haplogroup CT. For details on mutations see legend to Figure 1. Dashed lines indicate putative branchings (no positive control available). The position of V248 (haplogroup C2) and V87 (haplogroup C3) compared to mutations that define internal branches was not determined. Note that mutations V45, V69 and V88 have been previously mapped (Cruciani et al. 2010; Eur J Hum Genet 18∶800807).
Molecular Dissection of the Basal Clades in the Human Y Chromosome Phylogenetic Tree (2011)
quote:"haplogroup CF and DE molecular ancestors first evolved inside Africa and subsequently contributed as Y chromosome founders to pioneering migrations that successfully colonized Asia. While not proof, the DE and CF bifurcation (Figure 8d ) is consistent with independent colonization impulses possibly occurring in a short time interval."
Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations
--Peter A. Underhill , Toomas Kivisild - 2007
The Mal'ta boy didn't fell from the sky, onto Siberia near Lake Baikal?
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
quote:Originally posted by the lioness,: Not all haplogroups originate in Africa even if they're are also found in Africa because it depends on where they started.
The origin of a haplogroup is strongly suggested by
a) high frequency b) high diversity c) DNA analysis of ancient human remains
Basal!
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
posted
If M269 is found in Central, South,North Africa and etc., it is widespread because it is not situated in just one part of the African Continent.
This supports my contention that this haplogroup is widespread in Africa.
LOL. Montellano the Kivisild articles makes your whole paper invalid and your arguments prima facie evidence of your lack of knowledge about Indian genetics. In the 1999 article, Kivisild admits that Dravidians carry M1, which is an African haplogroup.
This article is just as relevant today as it was in 1999. People carry a particular gene do not just disappear.
The Eastern African hg M1, HVS-I signature motif is 16,129, 16,189, 16,223, 16,249, and 16,311. In the Kivisild et al figure below we see the same motif. The mutations are shown less 16,000.
Here you can clearly see:mutations 129,189, 223 and 311, in Indian M1, in Figure 3, of Kivisild et al, 1999.
In the Kivisild et al 1999 study of Indian mtDNA around 15% carried haplogroup M1. See:
In Table 4, Kivisild et al, 1999, we see the frequency of M1 in India.There are 217 million Dravidian speakers in India, if we compare the frequency of M1 carriers to the Dravidian speaking community around 32 million people carry M1.
The frequency of 15% of the Dravidians carrying M1 shows the presence of M1 in India. As a result, your arguments are false and invalid. .
Posts: 13012 | From: Chicago | Registered: Jan 2006
| IP: Logged |
quote:Originally posted by Troll Patrol # Ish Gebor:
quote:Originally posted by Quetzalcoatl:
quote:Originally posted by Clyde Winters:
In the Table I cite the source and the frequency. Only a fool and moron would accept your statements as valid given the fact you have repeatedly shown that the figures are correct.
Stop trying to steal the history of Black people.
You can whirl , you can spin, you can emit lots of spam BUT there is no R-M269 in Africa, R-V88 (R1b1) is only present in a limited area in Africa, there is no R-M173 (R1) in Africa. There is nothing to refute. Something has to proven before it can be refuted. It is up to the proposer to produce the evidence for a proposal. You have only produced misquotes. Please quote word-for-word refereed papers that say "M-269 is widespread in Africa" "M-173 is widespread in Africa, "R=P25*(R1b1) is widespread in Africa.
quote: The Y chromosome Alu polymorphism (YAP, also called M1) defines the deep-rooted haplogroup D/E of the global Y-chromosome phylogeny [1]. This D/E haplogroup is further branched into three sub-haplogroups DE*, D and E (Figure 1). The distribution of the D/E haplogroup is highly regional, and the three subgroups are geographically restricted to certain areas, therefore informative in tracing human prehistory (Table 1). The sub-haplogroup DE*, presumably the most ancient lineage of the D/E haplogroup was only found in Africans from Nigeria [2], supporting the "Out of Africa" hypothesis about modern human origin. The sub-haplogroup E (E-M40), defined by M40/SRY4064 and M96, was also suggested originated in Africa [3-6], and later dispersed to Middle East and Europe about 20,000 years ago [3,4]. Interestingly, the sub-haplogroup D defined by M174 (D-M174) is East Asian specific with abundant appearance in Tibetan and Japanese (3040%), but rare in most of other East Asian populations and populations from regions bordering East Asia (Central Asia, North Asia and Middle East) (usually less than 5%) [5-7]. Under D-M174, Japanese belongs to a separate sub-lineage defined by several mutations (e.g. M55, M57 and M64 etc.), which is different from those in Tibetans implicating relatively deep divergence between them [1]. The fragmented distribution of D-M174 in East Asia seems not consistent with the pattern of other East Asian specific lineages, i.e. O3-M122, O1-M119 and O2-M95 under haplogroup O [8,9].
quote: Further refinement awaits the finding of new markers especially within paragroup E3a*-M2. The microsatellite profile of the DE* individual is one mutational step away from the allelic state described for Nigerians (DYS390*21, DYS388 not tested; [37], therefore suggesting a common ancestry but not elucidating the phylogenetics.
Haplogroup DE* in Guinea-Bissau:
Y-chromosomal diversity in the population of Guinea-Bissau: a multiethnic perspective
quote: There has been considerable debate on the geographic origin of the human Y chromosome Alu polymor- phism (YAP). Here we report a new, very rare deep-rooting haplogroup within the YAP clade, together with data on other deep-rooting YAP clades. The new haplogroup, found so far in only five Nigerians, is the least-derived YAP haplogroup according to currently known binary markers. However, because the interior branching order of the Y chromosome genealogical tree remains unknown, it is impossible to impute the origin of the YAP clade with certainty. We discuss the problems presented by rare deep-rooting lineages for Y chromosome phylogeography.
Haplogroup DE* in Nigerians:
Rare Deep-Rooting Y Chromosome Lineages in Humans: Lessons for Phylogeography
Y-DNA haplogroup R-M207 is believed to have arisen approximately 27,000 years ago in Asia. The two currently defined subclades are R1 and R2.
quote:Out of Africa haplogroups.
All Y-clades that are not exclusively African belong to the macro-haplogroup CT, which is defined by mutations M168, M294 and P9.1 [14,31] and is subdivided into two major clades, DE and CF [1,14].
In a recent study [16], sequencing of two chromosomes belonging to haplogroups C and R, led to the identification of 25 new mutations, eleven of which were in the C-chromosome and seven in the R-chromosome.
Here, the seven mutations which were found to be shared by chromosomes of haplogroups C and R [16], were also found to be present in one DE sample (sample 33 in Table S1), and positioned at the root of macro-haplogroup CT (Figure 1 and Figure S1)
Structure of the macro-haplogroup CT. For details on mutations see legend to Figure 1. Dashed lines indicate putative branchings (no positive control available). The position of V248 (haplogroup C2) and V87 (haplogroup C3) compared to mutations that define internal branches was not determined. Note that mutations V45, V69 and V88 have been previously mapped (Cruciani et al. 2010; Eur J Hum Genet 18∶800807).
Molecular Dissection of the Basal Clades in the Human Y Chromosome Phylogenetic Tree (2011)
quote:"haplogroup CF and DE molecular ancestors first evolved inside Africa and subsequently contributed as Y chromosome founders to pioneering migrations that successfully colonized Asia. While not proof, the DE and CF bifurcation (Figure 8d ) is consistent with independent colonization impulses possibly occurring in a short time interval."
Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations
--Peter A. Underhill , Toomas Kivisild - 2007
The Mal'ta boy didn't fell from the sky, onto Siberia near Lake Baikal?
Correct. Geneticists like to look at haplogroups in isolation. The archaeology makes it clear that the first Europeans came from Africa and they were Khoisan, because the same culutral traditions of Cro-Magnon man in Europe were found in Southern Africa. It is clear that Mal'ta man's population did not just fall from the sky.
Posts: 13012 | From: Chicago | Registered: Jan 2006
| IP: Logged |
quote:Originally posted by the lioness,: Not all haplogroups originate in Africa even if they're are also found in Africa because it depends on where they started.
The origin of a haplogroup is strongly suggested by
a) high frequency b) high diversity c) DNA analysis of ancient human remains
High frequency and diversity can not tell us anything about origin because these features are are associated with contemporary populations. Since they relate only to contemporary populations, the story they tell is about the people who presently live in the area. These markers have little validity, because the contemporary population may not represent the original inhabitant of a particular region. This is especially true of European whites who only recently settled Europe, after they migrated from Central Asia.
posted
Klyosov in criticizing my article exagerates what I wrote. He implies that I was trying to decieve the readers about the frequency of R1 in Africa. This is false.
I specifically stated the frequency of R1 among African populations throughout my 2011 paper.
quote:
Y-chromosome R1 is found throghout Africa. The pristine form of R1-M173 is only found in Africa (Coia et al, 2005; Cruciani et al, 2002, 2010). The age of ychromosome R is 27ky. Most researchers believe that R(M173) is 18.5 ky.There is a great diversity of the macrohaplogroup R in Africa (See Figure 1). Ychromosome R is characterized by M207/V45. The V45 mutation is found among African populations ( Cruciani et al ,2010). ISOGG 2010 Y-DNA haplogroup tree makes it clear that V45 is phylogenetically equivalent to M207.The most common R haplogroup in Africa is R1 (M173). The predominant haplogroup is R1b (Berniell-Lee et al,2009; Coia et al, 2005; Winters, 2010b; Wood et al, 2009). Cruciani et al (2010) discovered new R1b mutations including V7, V8, V45, V69, and V88. Geography appears to play a significant role in the distribution of haplogroup R in Africa. Cruciani et al (2010) has renamed the R*- M173 (R P-25) in Africa V88. The TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010). Y-chromosome V88 (R1b1a) has its highest frequency among Chadic speakers, while the carriers of V88 among Niger-Congo speakers (predominately Bantu people) range between 2-66% ( Cruciani et al, 2010; Bernielle-Lee et al, 2009). Haplogroup V88 includes the mutations M18, V35 and V7. Cruciani et al (2010) revealed that R-V88 is also carried by Eurasians including the distinctive mutations M18, V35 and V7. R1b1-P25 is found in Western Eurasia. Haplogroup R1b1* is found in Africa at various frequencies. BerniellLee et al (2009) found in their study that 5.2% carried Rb1*. The frequency of R1b1* among the Bantu ranged from 2-20. The bearers of R1b1* among the Pygmy populations ranged from 1-25% (Berniell-Lee et al, 2009). The frequency of R1b1 among Guinea-Bissau populations was 12% (Carvalho et al,2010).
As you can see I did admit that only 5.2% of the pygmy carried R1.
In relation to R-M269 in Africa I wrote:
quote: Around 0.1 of Sub Saharan Africans carry R1b1b2. Wood et al (2009) found that Khoisan (2.2%) and Niger-Congo (0.4%) speakers carried the R-M269 y-chromosome. The Khoisan also carry RM343 (R1b) and M 198 (R1a1) (Naidoo et al., 2010) the archaeological and linguistic data indicate the successful colonization of Asia by SubSaharan Africans from Nubia 5-4kya (Winters, 2007,2008, 2010c). The archaeological evidence makes it clear that around 4kya intercultural style artifacts connected Africa and Eurasia (Winters, 2007,2010c).
As a result, I did not attempt to decieve anyone about the frequency of R1 in Africa as the author implies.
In fact recent research on y-haplogroups in Africa suggest that R1-M269 is also widespread in Africa.
Above is a figure from Gonzalez et al. The Gonzalez et al article found that 10 out of 19 subjects in the study carried R1b1-P25 or M269. This is highly significant because it indicates that 53% of the R1 carriers in this study were M269, this finding is further proof of the widespread nature of this so-called Eurasian genes in Africa among populations that have not mated with Europeans.
The R1 haplogroup probably originated in Africa.
.
.
. The phylogeography of R1 in Africa makes it clear that this y-chromosome is spread globally across Africa and includes the genetic structure of diverse African populations including Berber, Chadic, Cushitic, Khoisan,Pygmy, Niger-Congo, Nilo-Saharan and Semitic speaking African populations (Berniell-Lee et al, 2009; Cruciani et al, 2010; Wood et al, 2009). The fact that Dravidians carry the R haplogroup illustrate the recent introduction of R y-chromosome to Eurasia.
Abu-Amero et al (20009) reveal the fact that Dravidians carry the R haplogroups illustrate the recent introduction of Ry-chromosomes to Eurasia. The frequency of haplotype M173 in Eurasia is as follows: Anatolia 0.19%, Iran 2.67%, Iraq 0.49% Oman 1.0%, Pakistan 0.57% and Oman 1.0% . This contrast sharply with the widespread distribution of R1 in Africa that ranges between 7- 95% in various parts of Africa, especially Cameroon (Coia et al, 2005). Coia et al (2005) has revealed that no maternal Eurasian lineages have been found among Sub-Saharan Africans with a R1- M173 profile. Haplogroup V88 has the greatest frequency in Africa. It is predominately carried by Chadic speakers, ranges between 2-60% among Central African Niger-Congo speakers (Cruciani et al, 2010). Researchers have found that the TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010).
The vast majority of Africans belong to the y-chromosome E macrohaplogroup. Phylogenetically haplogroup R1b is mainly found in West Africa and the Sahel.
In this region the frequency of R-M173 can range between 85-100% among some Niger Congo speakers in Cameroon (Cruciani et al, 2010). The paternal record of M173 on the African continent illustrates a greater distribution of this y-chromosome among varied African populations than, in Asia.
The greatest diversity of R1b in Africa is highly suggestive of an Africa origin for this male lineage because it is not isolated to just one part of Africa.
Archaeological (Lal, 1963), genetic (Winters, 2008;2010a), placenames (Balakrishnan, 2005) and linguistic data group (Aravanan,1979,1980; Upadhyaya, 1976,1979; Winters 1985a,1985b, 1989) linking Africans and Dravidian support the recent demic diffusion of SubSaharan Africans and gene flow from Africa to Eurasia. An early colonization of Eurasia 4kya by Sub-Saharan Africans and Dravidian carriers of R1-M173 is the best scenario to explain the high frequency and widespread geographical distribution of this y-chromosome on the African continent (Winters, 2010c). Given the greatest diversity of R1- M173, this is the most parsimonious model explaining the frequency of R-M173 in Africa.
In conclusion, the R macrohaplogroup probably originated in Africa. In my paper POSSIBLE AFRICAN ORIGIN OF Y-CHROMOSOME R1-M173 , I argue that the P clade originated in Africa because 1) the age of R-V88 and 2) the widespread nature of R1 in Africa.
Researchers have found that the TMRCA of V88 was 9200-5600 kya (Cruciani et al, 2010). Eurasians carry the M269 (R1b1b2) mutation. The subclades of R1b1b2 include Rh1b1b2g (U106) (TMRCA 8.3kya) and R1b1b2h (U152) (TMRCA 7.4kya). The most recent common ancestor for R1b1b2 is probably 8kya (Balaresque et al, 2010).
In Africa we find R-M269 and V88. Clearly, R-V88 is older than R-M269 there is no evidence of archaeological evidence of a back migration or haplogroup R into Africa, but there is evidence of the migration of the Kushites and Proto-Saharans into Eurasia from Middle Africa. This supports the proposition the R haplogroups originated in Africa, not Eurasia.
Posts: 13012 | From: Chicago | Registered: Jan 2006
| IP: Logged |
LOL. Montellano the Kivisild articles makes your whole paper invalid and your arguments prima facie evidence of your lack of knowledge about Indian genetics. In the 1999 article, Kivisild admits that Dravidians carry M1, which is an African haplogroup.
This article is just as relevant today as it was in 1999. People carry a particular gene do not just disappear.
The Eastern African hg M1, HVS-I signature motif is 16,129, 16,189, 16,223, 16,249, and 16,311. In the Kivisild et al figure below we see the same motif. The mutations are shown less 16,000.
Here you can clearly see:mutations 129,189, 223 and 311, in Indian M1, in Figure 3, of Kivisild et al, 1999.
In the Kivisild et al 1999 study of Indian mtDNA around 15% carried haplogroup M1. See:
In Table 4, Kivisild et al, 1999, we see the frequency of M1 in India.There are 217 million Dravidian speakers in India, if we compare the frequency of M1 carriers to the Dravidian speaking community around 32 million people carry M1.
The frequency of 15% of the Dravidians carrying M1 shows the presence of M1 in India. As a result, your arguments are false and invalid.
Predictably, the air wavs are now filled with your irrelevant spam.
Kivisild 1999 is, and has been for years, incorrect because the nomenclature for the haplogroups have changed . as I point out in my paper the haplogroup called M1 by Kivisild in 1999-- has, for years been called M3 by everybody but you. Anyone can check this out in phylotree- the official nomenclature site.
Posts: 833 | From: Austin, TX | Registered: Jan 2007
| IP: Logged |
quote:Originally posted by Clyde Winters: [LOL. Montellano the Kivisild articles makes your whole paper invalid and your arguments prima facie evidence of your lack of knowledge about Indian genetics. In the 1999 article, Kivisild admits that Dravidians carry M1, which is an African haplogroup.
This article is just as relevant today as it was in 1999. People carry a particular gene do not just disappear.
.
The 1999 paper is invalid. The people did not disappear, however the name of their haplotype changed (by the committee in charge of nomenclature) to M3.
quote:The Eastern African hg M1, HVS-I signature motif is 16,129, 16,189, 16,223, 16,249, and 16,311. In the Kivisild et al figure below we see the same motif. The mutations are shown less 16,000.
quote:In Table 4, Kivisild et al, 1999, we see the frequency of M1 in India.There are 217 million Dravidian speakers in India, if we compare the frequency of M1 carriers to the Dravidian speaking community around 32 million people carry M1. [QB][/ The frequency of 15% of the Dravidians carrying M1 shows the presence of M1 in India. As a result, your arguments are false and invalid.
NO, 23 million people in India carry M3. Get over it!!
Posts: 833 | From: Austin, TX | Registered: Jan 2007
| IP: Logged |
quote:Originally posted by Troll Patrol # Ish Gebor:
quote:Originally posted by Quetzalcoatl:
quote:Originally posted by Clyde Winters:
In the Table I cite the source and the frequency. Only a fool and moron would accept your statements as valid given the fact you have repeatedly shown that the figures are correct.
Stop trying to steal the history of Black people.
You can whirl , you can spin, you can emit lots of spam BUT there is no R-M269 in Africa, R-V88 (R1b1) is only present in a limited area in Africa, there is no R-M173 (R1) in Africa. There is nothing to refute. Something has to proven before it can be refuted. It is up to the proposer to produce the evidence for a proposal. You have only produced misquotes. Please quote word-for-word refereed papers that say "M-269 is widespread in Africa" "M-173 is widespread in Africa, "R=P25*(R1b1) is widespread in Africa.
quote: The Y chromosome Alu polymorphism (YAP, also called M1) defines the deep-rooted haplogroup D/E of the global Y-chromosome phylogeny [1]. This D/E haplogroup is further branched into three sub-haplogroups DE*, D and E (Figure 1). The distribution of the D/E haplogroup is highly regional, and the three subgroups are geographically restricted to certain areas, therefore informative in tracing human prehistory (Table 1). The sub-haplogroup DE*, presumably the most ancient lineage of the D/E haplogroup was only found in Africans from Nigeria [2], supporting the "Out of Africa" hypothesis about modern human origin. The sub-haplogroup E (E-M40), defined by M40/SRY4064 and M96, was also suggested originated in Africa [3-6], and later dispersed to Middle East and Europe about 20,000 years ago [3,4]. Interestingly, the sub-haplogroup D defined by M174 (D-M174) is East Asian specific with abundant appearance in Tibetan and Japanese (3040%), but rare in most of other East Asian populations and populations from regions bordering East Asia (Central Asia, North Asia and Middle East) (usually less than 5%) [5-7]. Under D-M174, Japanese belongs to a separate sub-lineage defined by several mutations (e.g. M55, M57 and M64 etc.), which is different from those in Tibetans implicating relatively deep divergence between them [1]. The fragmented distribution of D-M174 in East Asia seems not consistent with the pattern of other East Asian specific lineages, i.e. O3-M122, O1-M119 and O2-M95 under haplogroup O [8,9].
quote: Further refinement awaits the finding of new markers especially within paragroup E3a*-M2. The microsatellite profile of the DE* individual is one mutational step away from the allelic state described for Nigerians (DYS390*21, DYS388 not tested; [37], therefore suggesting a common ancestry but not elucidating the phylogenetics.
Haplogroup DE* in Guinea-Bissau:
Y-chromosomal diversity in the population of Guinea-Bissau: a multiethnic perspective
quote: There has been considerable debate on the geographic origin of the human Y chromosome Alu polymor- phism (YAP). Here we report a new, very rare deep-rooting haplogroup within the YAP clade, together with data on other deep-rooting YAP clades. The new haplogroup, found so far in only five Nigerians, is the least-derived YAP haplogroup according to currently known binary markers. However, because the interior branching order of the Y chromosome genealogical tree remains unknown, it is impossible to impute the origin of the YAP clade with certainty. We discuss the problems presented by rare deep-rooting lineages for Y chromosome phylogeography.
Haplogroup DE* in Nigerians:
Rare Deep-Rooting Y Chromosome Lineages in Humans: Lessons for Phylogeography
Y-DNA haplogroup R-M207 is believed to have arisen approximately 27,000 years ago in Asia. The two currently defined subclades are R1 and R2.
quote:Out of Africa haplogroups.
All Y-clades that are not exclusively African belong to the macro-haplogroup CT, which is defined by mutations M168, M294 and P9.1 [14,31] and is subdivided into two major clades, DE and CF [1,14].
In a recent study [16], sequencing of two chromosomes belonging to haplogroups C and R, led to the identification of 25 new mutations, eleven of which were in the C-chromosome and seven in the R-chromosome.
Here, the seven mutations which were found to be shared by chromosomes of haplogroups C and R [16], were also found to be present in one DE sample (sample 33 in Table S1), and positioned at the root of macro-haplogroup CT (Figure 1 and Figure S1)
Structure of the macro-haplogroup CT. For details on mutations see legend to Figure 1. Dashed lines indicate putative branchings (no positive control available). The position of V248 (haplogroup C2) and V87 (haplogroup C3) compared to mutations that define internal branches was not determined. Note that mutations V45, V69 and V88 have been previously mapped (Cruciani et al. 2010; Eur J Hum Genet 18∶800807).
Molecular Dissection of the Basal Clades in the Human Y Chromosome Phylogenetic Tree (2011)
quote:"haplogroup CF and DE molecular ancestors first evolved inside Africa and subsequently contributed as Y chromosome founders to pioneering migrations that successfully colonized Asia. While not proof, the DE and CF bifurcation (Figure 8d ) is consistent with independent colonization impulses possibly occurring in a short time interval."
Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations
--Peter A. Underhill , Toomas Kivisild - 2007
The Mal'ta boy didn't fell from the sky, onto Siberia near Lake Baikal?
Correct. Geneticists like to look at haplogroups in isolation. The archaeology makes it clear that the first Europeans came from Africa and they were Khoisan, because the same culutral traditions of Cro-Magnon man in Europe were found in Southern Africa. It is clear that Mal'ta man's population did not just fall from the sky.
Yep,
quote:"No southwest Asian specific clades for M1 or U6 were discovered. U6 and M1 frequencies in North Africa, the Middle East and Europe DO NOT FOLLOW similar patterns, and their sub-clade divisions do not appear to be compatible with their shared history reaching back to the Early Upper Palaeolithic."
--Erwan Pennarun, Toomas Kivisild
(2012) Divorcing the Late Upper Palaeolithic demographic histories of mtDNA haplogroups M1 and U6 in Africa
quote:... the M1 presence in the Arabian peninsula signals a predominant East African influence since the Neolithic onwards.
-- Petraglia, M and Rose, J
(2010). The Evolution of Human Populations in Arabia:
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
quote:Originally posted by Clyde Winters: [LOL. Montellano the Kivisild articles makes your whole paper invalid and your arguments prima facie evidence of your lack of knowledge about Indian genetics. In the 1999 article, Kivisild admits that Dravidians carry M1, which is an African haplogroup.
This article is just as relevant today as it was in 1999. People carry a particular gene do not just disappear.
.
The 1999 paper is invalid. The people did not disappear, however the name of their haplotype changed (by the committee in charge of nomenclature) to M3.
quote:The Eastern African hg M1, HVS-I signature motif is 16,129, 16,189, 16,223, 16,249, and 16,311. In the Kivisild et al figure below we see the same motif. The mutations are shown less 16,000.
quote:In Table 4, Kivisild et al, 1999, we see the frequency of M1 in India.There are 217 million Dravidian speakers in India, if we compare the frequency of M1 carriers to the Dravidian speaking community around 32 million people carry M1. [QB][/ The frequency of 15% of the Dravidians carrying M1 shows the presence of M1 in India. As a result, your arguments are false and invalid.
NO, 23 million people in India carry M3. Get over it!!
LOL. How could haplogroup M1 change to M3, when Kivisild presents both haplohroups in Figure 3.
Abu-Amero et al (2009) reveal the fact that Dravidians carry the R haplogroups illustrate the recent introduction of R y-chromosomes to Eurasia. The frequency of haplotype M173 in Eurasia is as follows: Anatolia 0.19%, Iran 2.67%, Iraq 0.49% Oman 1.0%, Pakistan 0.57% and Oman 1.0% . This contrast sharply with the widespread distribution of R1 in Africa that ranges between 7- 95% in various parts of Africa,
Abu-Amero's 2009 article is called Saudi Arabian Y-Chromosome diversity and its relationship with with nearby regions.
The regions discussed in the article are far from being the whole of Eurasia
Saudi Arabia Qatar UAE Oman Yemen Somalia Egypt Jordan Iraq Anataolia Iran Pakistan Lebanon
Why are you saying that the article reveals the fact that Dravidians carry the R haplogroups when the article says nothing about Dravidians ?
Why are you saying the frequency of haplotype M173 in Eurasia is represented by the above countries when the above countries are only a particular region of Eurasia and are NOT the countries with the HIGH frequencies of M173 ?
________________________________
As for M17 the descendant of M173 in India it is found at the higher frequencies amoung the Indo European speakers, the Hindi spekaing Brahmin and at lower frequencies in the Dravidians And all of these people of course also Eurasians
An it is also found in high frequenices in Central Asians and some Eastern Europeans
You like to speak about the Aryan invasions of India. That of course will also have a genetic marker
We are talking about R1a here
Of course in Cameroon it's a clade of R1b not R1a
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
Abu-Amero et al (2009) reveal the fact that Dravidians carry the R haplogroups illustrate the recent introduction of R y-chromosomes to Eurasia. The frequency of haplotype M173 in Eurasia is as follows: Anatolia 0.19%, Iran 2.67%, Iraq 0.49% Oman 1.0%, Pakistan 0.57% and Oman 1.0% . This contrast sharply with the widespread distribution of R1 in Africa that ranges between 7- 95% in various parts of Africa,
Abu-Amero's 2009 article is called Saudi Arabian Y-Chromosome diversity and its relationship with with nearby regions.
The regions discussed in the article are far from being the whole of Eurasia
Saudi Arabia Qatar UAE Oman Yemen Somalia Egypt Jordan Iraq Anataolia Iran Pakistan Lebanon
Why are you saying that the article reveals the fact that Dravidians carry the R haplogroups when the article says nothing about Dravidians ?
Why are you saying the frequency of haplotype M173 in Eurasia is represented by the above countries when the above countries are only a particular region of Eurasia and are NOT the countries with the HIGH frequencies of M173 ?
________________________________
As for M17 the descendant of M173 in India it is found at the higher frequencies amoung the Indo European speakers, the Hindi spekaing Brahmin and at lower frequencies in the Dravidians And all of these people of course also Eurasians
An it is also found in high frequenices in Central Asians and some Eastern Europeans
You like to speak about the Aryan invasions of India. That of course will also have a genetic marker
We are talking about R1a here
Of course in Cameroon it's a clade of R1b not R1a
These questions have nothing to do with this thread. We are talking about M173 in Africa and M1 in India.
Stay on topic .
Posts: 13012 | From: Chicago | Registered: Jan 2006
| IP: Logged |
Abu-Amero et al (2009) reveal the fact that Dravidians carry the R haplogroups illustrate the recent introduction of R y-chromosomes to Eurasia. The frequency of haplotype M173 in Eurasia is as follows: Anatolia 0.19%, Iran 2.67%, Iraq 0.49% Oman 1.0%, Pakistan 0.57% and Oman 1.0% . This contrast sharply with the widespread distribution of R1 in Africa that ranges between 7- 95% in various parts of Africa,
Abu-Amero's 2009 article is called Saudi Arabian Y-Chromosome diversity and its relationship with with nearby regions.
The regions discussed in the article are far from being the whole of Eurasia
Saudi Arabia Qatar UAE Oman Yemen Somalia Egypt Jordan Iraq Anataolia Iran Pakistan Lebanon
Why are you saying that the article reveals the fact that Dravidians carry the R haplogroups when the article says nothing about Dravidians ?
Why are you saying the frequency of haplotype M173 in Eurasia is represented by the above countries when the above countries are only a particular region of Eurasia and are NOT the countries with the HIGH frequencies of M173 ?
________________________________
As for M17 the descendant of M173 in India it is found at the higher frequencies amoung the Indo European speakers, the Hindi spekaing Brahmin and at lower frequencies in the Dravidians And all of these people of course also Eurasians
An it is also found in high frequenices in Central Asians and some Eastern Europeans
You like to speak about the Aryan invasions of India. That of course will also have a genetic marker
We are talking about R1a here
Of course in Cameroon it's a clade of R1b not R1a
These questions have nothing to do with this thread. We are talking about M173 in Africa and M1 in India.
Stay on topic .
Clyde I can't believe you don't know this, haplogroup R-M17 is the sub clade of M173 found in India. But to the point you quoted Abu-Amero in this thread and apparently didn't know what you were talking about And you don't understand that R1 has two major clades R1a and R1b. Clyde you don't even know the basics, haplgroup M17 is Y dna, short for R-M17 aka R1a1* and has nothing to do with M1 which is mtDNA The one who was off topic was you, talking about an article on Saudi Arabia and nearby regions and pretending that represented frequencies of Eurasia in general
In case you've forgoten you brought up this topic in this thread
Again >
quote:Originally posted by Clyde Winters:
Abu-Amero et al (2009) reveal the fact that Dravidians carry the R haplogroups illustrate the recent introduction of R y-chromosomes to Eurasia. The frequency of haplotype M173 in Eurasia is as follows: Anatolia 0.19%, Iran 2.67%, Iraq 0.49% Oman 1.0%, Pakistan 0.57% and Oman 1.0% . This contrast sharply with the widespread distribution of R1 in Africa that ranges between 7- 95% in various parts of Africa,
^^^ So if you are going to make very misleading statements like this don't expect it to go unaddressed
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
quote:An independent high resolution MSY phylogeny has been recently obtained from 2,870 Y-SNPs discovered (or re-discovered) in the course of a large whole-genome re-sequencing study, but the observed variable sites all belong to the recent out of Africa CT clade [15]. Recently, in a re-sequencing study of the Y chromosome, the root of the tree moved to a new position and several changes at the basal nodes of the phylogeny were introduced [16]. Interestingly, the estimated coalescence age and deep branching pattern of the revised MSY tree appear to be more similar to those of the mtDNA phylogeny [17],[18] than previously reported [1].
[...]
Three of the seven R-specific mutations (V45, V69 and V88) were previously mapped within haplogroup R [34], whereas the remaining four mutations have been here positioned at the root of haplogroups F (V186 and V205), K (V104) and P (V231)(Figure S1) through the analysis of 12 haplogroup F samples (samples 4051, in Table S1).
[...]
All Y-clades that are not exclusively African belong to the macro-haplogroup CT, which is defined by mutations M168, M294 and P9.1 [14],[31] and is subdivided into two major clades, DE and CF [1],[14]. In a recent study [16], sequencing of two chromosomes belonging to haplogroups C and R, led to the identification of 25 new mutations, eleven of which were in the C-chromosome and seven in the R-chromosome. Here, the seven mutations which were found to be shared by chromosomes of haplogroups C and R [16]
[...]
Two A1b chromosomes from a previous work (one from Algeria and one from Cameroon)[16] were included in this study together with two newly identified A1b chromosomes, whose geographic origin can be traced back to west-central Africa (Ghana) on the basis of the microsatellite profile (data not shown).
Structure of the macro-haplogroup CT. For details on mutations see legend to Figure 1. Dashed lines indicate putative branchings (no positive control available). The position of V248 (haplogroup C2) and V87 (haplogroup C3) compared to mutations that define internal branches was not determined. Note that mutations V45, V69 and V88 have been previously mapped (Cruciani et al. 2010; Eur J Hum Genet 18∶800807).
--Scozzari R, Massaia A, DAtanasio E, Myres NM, Perego UA, et al.(2012) Molecular Dissection of the Basal Clades in the Human Y Chromosome Phylogenetic Tree. PLoS ONE 7(11): e49170. doi:10.1371/journal.pone.00491 70
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
Abu-Amero et al (2009) reveal the fact that Dravidians carry the R haplogroups illustrate the recent introduction of R y-chromosomes to Eurasia. The frequency of haplotype M173 in Eurasia is as follows: Anatolia 0.19%, Iran 2.67%, Iraq 0.49% Oman 1.0%, Pakistan 0.57% and Oman 1.0% . This contrast sharply with the widespread distribution of R1 in Africa that ranges between 7- 95% in various parts of Africa,
^^^ So if you are going to make very misleading statements like this don't expect it to go unaddressed
This is irrelevant because I cite the source for the frequency of haplotype M173 in Eurasia, which are much lower than those in Africa. These figures are not mine.
I have not discussed R1a, because it relates to the Dravidian speakers who originated in Africa. As a result, since they took this haplogroup to India it also has to be of African origin so there is no need to discuss this issue.
Researchers like to claim that R1a, is not found among Africans but this is false as seen in this Figure from Berniell-Lee . . . LOL. Your problem is that you can't believe that much of the population genetics literature is bs. It is bs because the researchers only present statistics and frequencies they already agree with,à la Bayesian statistics.
You can try to find some minutiae detail and try to use it to support your proposition but you will fail because I cited the proper figures. Montellano has not proven anything in this thread. In my 2011 article I cite reliable figures for M269 in Africa. It is found in North, South, East and West Africa, just like V88, so the haplogroup is widespread.
Also, in 1999, Kivisild reported that 15% of the Dravidians or 32 million people carried haplogroup M1, and these authors also cited figures for M3. If M1 and M3 already existed in 1999, there is no way you can later say M1 was really M3 when they already illustrated in Figure 3, that both haoplogroups were present in India. .
. I only entertain this issue because I like to show what great deceivers you and Montellano are.
Abu-Amero et al (2009) reveal the fact that Dravidians carry the R haplogroups illustrate the recent introduction of R y-chromosomes to Eurasia. The frequency of haplotype M173 in Eurasia is as follows: Anatolia 0.19%, Iran 2.67%, Iraq 0.49% Oman 1.0%, Pakistan 0.57% and Oman 1.0% . This contrast sharply with the widespread distribution of R1 in Africa that ranges between 7- 95% in various parts of Africa,
^^^ So if you are going to make very misleading statements like this don't expect it to go unaddressed
This is irrelevant because I cite the source for the frequency of haplotype M173 in Eurasia, which are much lower than those in Africa. These figures are not mine.
You quoted a source on frequencies of R1 in Saudi Arabia and nearby countries that have low frequencies of R1 and said that represented Eurasia Meanwhile you left out R1 frequencies of Europe, India and Central Asia. Everybody knows what you did
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
Abu-Amero et al (2009) reveal the fact that Dravidians carry the R haplogroups illustrate the recent introduction of R y-chromosomes to Eurasia. The frequency of haplotype M173 in Eurasia is as follows: Anatolia 0.19%, Iran 2.67%, Iraq 0.49% Oman 1.0%, Pakistan 0.57% and Oman 1.0% . This contrast sharply with the widespread distribution of R1 in Africa that ranges between 7- 95% in various parts of Africa,
^^^ So if you are going to make very misleading statements like this don't expect it to go unaddressed
This is irrelevant because I cite the source for the frequency of haplotype M173 in Eurasia, which are much lower than those in Africa. These figures are not mine.
You quoted a source on frequencies of R1 in Saudi Arabia and nearby countries that have low frequencies of R1 and said that represented Eurasia Meanwhile you left out R1 frequencies of Europe, India and Central Asia. Everybody knows what you did
Here, the seven mutations which were found to be shared by chromosomes of haplogroups C and R [16], were also found to be present in one DE sample (sample 33 in Table S1), and positioned at the root of macro-haplogroup CT (Figure 1 and Figure S1)
posted
The idea that haplgroup M maybe have originated in East Africa is a legitiimate theory but may or may not be true since the horn is a coastal region and has higher frequencies of Eurasian haplogroups as compared to other parts of sub saharan Africa Of the various sub clades only M1 is found in Africa, in the horn. Ethiopians have the highest frequencies at about 20%. M is the single most common mtDNA haplogroup in Asia and in the world is most diverse in India.
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
quote:Originally posted by the lioness,: The idea that haplgroup M maybe have originated in East Africa is a legitiimate theory but may or may not be true since the horn is a coastal region and has higher frequencies of Eurasian haplogroups as compared to other parts of sub saharan Africa Of the various sub clades only M1 is found in Africa, in the horn. Ethiopians have the highest frequencies at about 20%. M is the single most common mtDNA haplogroup in Asia and in the world is most diverse in India.
quote:Originally posted by the lioness,: The idea that haplgroup M maybe have originated in East Africa is a legitiimate theory but may or may not be true since the horn is a coastal region and has higher frequencies of Eurasian haplogroups as compared to other parts of sub saharan Africa Of the various sub clades only M1 is found in Africa, in the horn. Ethiopians have the highest frequencies at about 20%. M is the single most common mtDNA haplogroup in Asia and in the world is most diverse in India.
Not again.
You best can see the Arabian Peninsula as a bridge, not as a time maschine where things change in an instance. A mutation takes generation. Perhaps many generation to fully adapt.
M # T489C, C10400T, T14783C, G15043A
quote:"The presence of M haplogroup in Ethiopia, named M1, led to the proposal that haplogroup M originated in eastern Africa, approximately 60,000 years ago, and was carried towards Asia [34].
Macrohaplogroup M is ubiquitous in India and covers more than 70 per cent of the Indian mtDNA lineages [28], [36][38]. Recent studies on complete mtDNA sequences (~187) tried to resolve the phylogeny of Indian macrohaplogroup M. As a result, M2, M3, M4, M5, M6 [28], [36], [39][40], M18, M25 [38], M30, [41], M31 [42], [24] M33, M34, M35, M36, M37, M38, M39, M40 [22], M41, M42 [43], M43 [23], [44], M45 [45], M48, M49, and M50 [46] haplogroups of M that was identified in India helped to a certain extent in understanding M genealogy in diversified Indian populations. In the above background, extensive sequencing of complete mtDNA of South Asia, particularly India, is essential for better understanding of the peopling of the non-African continents, and pathogenesis of diseases in various ethnic groups with different matrilineal backgrounds."
--Adimoolam Chandrasekar et al. 2009
Updating Phylogeny of Mitochondrial DNA Macrohaplogroup M in India: Dispersal of Modern Human in South Asian Corridor
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
quote:Originally posted by the lioness,: The idea that haplgroup M maybe have originated in East Africa is a legitiimate theory but may or may not be true since the horn is a coastal region and has higher frequencies of Eurasian haplogroups as compared to other parts of sub saharan Africa Of the various sub clades only M1 is found in Africa, in the horn. Ethiopians have the highest frequencies at about 20%. M is the single most common mtDNA haplogroup in Asia and in the world is most diverse in India.
That fact that a haplogroup is found in Africa doesn't means it originates there. ' Some haplogroups found in modern Africans are the result of migrations form Eurasia. People have been out of Africa for at least 70,000 years. During this time some haplogroups formed outside of Africa. Clyde can you name one to show you are rational?
he origin of a haplogroup is strongly suggested by
a) high frequency b) high diversity c) DNA analysis of ancient human remains
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
quote:Originally posted by the lioness,: The idea that haplgroup M maybe have originated in East Africa is a legitiimate theory but may or may not be true since the horn is a coastal region and has higher frequencies of Eurasian haplogroups as compared to other parts of sub saharan Africa Of the various sub clades only M1 is found in Africa, in the horn. Ethiopians have the highest frequencies at about 20%. M is the single most common mtDNA haplogroup in Asia and in the world is most diverse in India.
That fact that a haplogroup is found in Africa doesn't means it originates there. ' Some haplogroups found in modern Africans are the result of migrations form Eurasia. People have been out of Africa for at least 70,000 years. During this time some haplogroups formed outside of Africa. Clyde can you name one to show you are rational?
he origin of a haplogroup is strongly suggested by
a) high frequency b) high diversity c) DNA analysis of ancient human remains
This map shows the diversity of haplogroup M in Africa.
. Cite the archaeological evidence of a back migration from Eurasia into Africa.
The diversity of macrohaplogroup M in Africa support an African origin.
posted
Clyde, were you aware, there are about 3 million Indian people in Africa, spanning 12 countries
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
quote:Originally posted by the lioness,: Clyde, were you aware, there are about 3 million Inidan people in Africa, spanning 12 countries
The East Indians in Africa, are mainly from North India. They usually believe they are white and during the colonial times they supported Europeans.
Mongoloid Indians in America, have always been a minority because they were usually hunter-gather or nomads. Black Native Americans were farmers and produced abundant surplus food stocks.
That is why whites have never seen mongoloid Indians as a threat.
Mongoloid Indians in America, have always been a minority because they were usually hunter-gather or nomads. Black Native Americans were farmers and produced abundant surplus food stocks.
Clyde, stop making up this fake and divisive racial concept of "Black" vs "Mongoloid" native Americans, it's a bad thng you are spreading
Even Black people who claim to have Native American ancestry attend the pow wows without contriving these two separate races of native Americans
If the 15 million blacks in this country were not primarily African and instead were primarily Native American many many more would speak Native American language or at least know some words, practice the culture and Europeans would have remarked that Native Americans looked exactly like West Africans, it's a farce you are pulling. Not that some blacks in America don't have Native ancestry but you grossly exaggerte to absurd proportions- and to the detriment of Native Americans.
Posts: 42919 | From: , | Registered: Jan 2010
| IP: Logged |
I'm puzzled. Why do we have a phylogeny of mtDNA when we are discussing the Y-chromosome?
Posts: 833 | From: Austin, TX | Registered: Jan 2007
| IP: Logged |
Mongoloid Indians in America, have always been a minority because they were usually hunter-gather or nomads. Black Native Americans were farmers and produced abundant surplus food stocks.
Clyde, stop making up this fake and divisive racial concept of "Black" vs "Mongoloid" native Americans, it's a bad thng you are spreading
Even Black people who claim to have Native American ancestry attend the pow wows without contriving these two separate races of native Americans
If the 15 million blacks in this country were not primarily African and instead were primarily Native American many many more would speak Native American language or at least know some words, practice the culture and Europeans would have remarked that Native Americans looked exactly like West Africans, it's a farce you are pulling. Not that some blacks in America don't have Native ancestry but you grossly exaggerte to absurd proportions- and to the detriment of Native Americans.
Stupid! How could they continue speaking their Native language when the white man made them slaves or classified them as "free colored" and forced them to learn English.
In some cases, the Black Native Americans spoke African languages such as Mande and Akan. African Lexical items and numerals were collected by Europeans as evidenced in the Nanticoke numerals that were collected by J.C. Pyrlaeus that are Mande.
. . Moreover, Europeans did discribe Native Americans as looking like West Africans. For example, the Nanticokes were described by B.S Barton as very dark
Posts: 13012 | From: Chicago | Registered: Jan 2006
| IP: Logged |
I'm puzzled. Why do we have a phylogeny of mtDNA when we are discussing the Y-chromosome?
Are we? If so, then there has been major confusion from the start. I however, tapped in somewhere in the middle of the conversation, where mtDNA Hg M was mentioned.
But yeah, earlier on the conversation was on male ancestry.
quote: The Y chromosome Alu polymorphism (YAP, also called M1) defines the deep-rooted haplogroup D/E of the global Y-chromosome phylogeny [1]. This D/E haplogroup is further branched into three sub-haplogroups DE*, D and E (Figure 1). The distribution of the D/E haplogroup is highly regional, and the three subgroups are geographically restricted to certain areas, therefore informative in tracing human prehistory (Table 1). The sub-haplogroup DE*, presumably the most ancient lineage of the D/E haplogroup was only found in Africans from Nigeria [2], supporting the "Out of Africa" hypothesis about modern human origin. The sub-haplogroup E (E-M40), defined by M40/SRY4064 and M96, was also suggested originated in Africa [3-6], and later dispersed to Middle East and Europe about 20,000 years ago [3,4]. Interestingly, the sub-haplogroup D defined by M174 (D-M174) is East Asian specific with abundant appearance in Tibetan and Japanese (3040%), but rare in most of other East Asian populations and populations from regions bordering East Asia (Central Asia, North Asia and Middle East) (usually less than 5%) [5-7]. Under D-M174, Japanese belongs to a separate sub-lineage defined by several mutations (e.g. M55, M57 and M64 etc.), which is different from those in Tibetans implicating relatively deep divergence between them [1]. The fragmented distribution of D-M174 in East Asia seems not consistent with the pattern of other East Asian specific lineages, i.e. O3-M122, O1-M119 and O2-M95 under haplogroup O [8,9].
quote:In the 1980s and 90s, improvements in the sequencing of DNA led to a number of researchers sequencing the mtDNA of numerous populations in an attempt to trace the evolution of humans and the way in which truly modern humans, Homo sapiens sapiens, populated Africa and spread out over the globe.
--Bernard Ortiz de Montellano
quote: Genetic genealogy is more within reach of the average person than ever, thanks to advances in sequencing technology that have helped the cost of genome sequencing dramatically plummet from nearly $3 billion in 2000 to near $1,000 nowadays. That sort of price reduction is mind-boggling, Ball said. Its as if, 15 years from now, I could get my own Mars rover.
quote:Originally posted by Troll Patrol # Ish Gebor:
quote:In the 1980s and 90s, improvements in the sequencing of DNA led to a number of researchers sequencing the mtDNA of numerous populations in an attempt to trace the evolution of humans and the way in which truly modern humans, Homo sapiens sapiens, populated Africa and spread out over the globe.
--Bernard Ortiz de Montellano
quote: Genetic genealogy is more within reach of the average person than ever, thanks to advances in sequencing technology that have helped the cost of genome sequencing dramatically plummet from nearly $3 billion in 2000 to near $1,000 nowadays. That sort of price reduction is mind-boggling, Ball said. Its as if, 15 years from now, I could get my own Mars rover.
Just as the cost of genetic sequencing has declined the mystery about what DNA can and can not prove has also become evident. In the past we thought that DNA could reliably tell us everything we needed to know about population movements and the history of mankind. As a result, when geneticists claimed this or that we assumed that what they were saying was true and valid.
We accepted this view because geneticists claimed they were based on statistics, and educated people have been conditioned to believe if you can explain a phenomena by giving it a statistic , the statistic can explain the phenomena. As a result, when geneticists used Bayesian statistic to explain the data they are researching, we assumed that the results are scientifically valid.
The problem with this assumption is that Bayesian statistics are not based on the scientific method. The scientific method demands that you have two groups. They are the control and experimental groups.An experimental group is the group in an experiment that receives the variable being tested. One variable is tested at a time. The experimental group is compared to a control group, which does not receive the test variable.
The scientific method is based on hypotheses testing. Hypotheses testing means that a researcher forms a hypothesis and test the hypothesis using a series of quantitative or qualitative statistical methods to determine the statistical significance of the hypothesis being tested. The scientific method is based on experimentation to test a hypothesis .
Population geneticists usually do not test hypotheses. They make inferences about data based on Bayesian statistical inferences. They do not use statistical methods to determine the statistical significance of a hypothesis, they use statistics to describe data being reviewed by the researcher based on the beliefs the researcher already holds about the data being reviewed..
Population genetics is a type of Expost facto research. Expost facto research design is a quasi-experimental type of study examining how an independent variable, present prior to the research study, affects a dependent variable.
Whereas the subjects in experimental research are randomly selected, the participants in Expost facto research , are not randomly selected or assigned.The genome of the research subjects is examined to determine the haplotypes and haplogroups carried by the participants in the study.
In population genetics research the researcher uses the Bayesian inference method of statistical inference. Bayesian statistical method, is a subjective research design/method that provides a rational method of updating the researcher's beliefs.
Since, the results of a Bayesian statistical analysis are a series of beliefs based on statistical inferences, the results can not stand alone. This is due to the reality, that any results, reported by a researcher are only a series of inferences based on the researchers belief about a phenomena backed up by a series statistical results. If the results are published without corresponding evidence from archaeology, anthropology, linguistics and or craniometrics the inferences are pure conjecture, because they reflect the attitudes already held by the researcher, confirmed by data selected by the researcher to support his or her beliefs.
There is a sociological basis behind how a researcher interprets data. Sociological research indicates that there are unconscious cognitive structures within each individual. Cognitive structures that hold the idealistic view of members of the academe that determine how they perceive "reality". These structures are called doxa.
Commenting on these schema Berlinerblau (1999) noted that "These types of theories share the assumption that human beings know things that they do not even know that they know; that they "possess" knowledge about the world which exists in some sort of cognitive substrate, beyond the realm of discourse" (p.106).Wacquant (1995) says that doxa is " a realm of implicit and unstated beliefs".
Given the research suggesting that doxa exist, support the view that some researchers allow their hatred of multiculturalism, ethnic prejudice and racism to define their discourse, teaching and writing about themes relating to groups " other" ,than their own cultural and ethnic group . Geneticist are human beings, and usually they bring their ethnic prejudices to their research.
The discovery that man originated in Africa and the first homo sapiens were Black caused a dilemma among white geneticists. They could no longer claim people were divided into races, so they stopped identifying people based on their race. Instead they knew that each race lived on a particular continent so they just gave the old races, new names whites "Western Eurasian", mongoloid people "Eastern Eurasian", and the negroes Sub-Saharan Africans.By applying the name Sub-Saharan African to the "negro" group, they made the rest of the negroes Polynesian/Melanesians and Australians.
Genetic sequencing was an exciting field in the beginning. Researchers reported their findings in journals. But a problem soon emerged. As researchers began to sequence non-European DNA, they soon discovered that Africans, were carrying the same haplogroups they had assigned to Eurasians.
This caused a dilemma because the genetic research was showing that everyone was just a African.Whites could not accept this so they began to check the names of haplogroups and do further sequencing of DNA to try and keep humanity divided into separate populations/races. Soon Eurasians who carried the M haplogroup, were now said to carry mtDNA haplogroup D, instead of M. This proved successful in identifying Eastern Eurasians and Polynesians.
But there were too many Dravidians carrying M1, which researchers said was an African haplogroup, so they had to erase M1 from India, the problem was that the literature on M haplogroups in India had been published in print media. And as a result, the evidence for M1 in India could never be totally erased.
They were able to keep up this deception for years; because establishment geneticist would repeat the new dogma in their publications. The same thing was done with the R haplogroup, which was just originally M173, and specific populations were divided into V88 for the Africans. and M269 for the whites.
The mythology surrounding mtDNA haplogroup M1, and y-Chromosomes V88 and M269, was stable until I came along. As I promoted the fact that Africans also carried M269, they changed the phylogeny of hg R, to begin with M207, but they forgot that up to 2011, V45 was classified as the same haplogroup as M207, which meant that Africans and Europeans were carrying the same haplogroups. So they changed the phylogeny of R, and made V45 distinct from M207.
The goal of lioness and Montellano is to use misdirection to maintain the myth Europeans--whites--carry a unique set of genes that separate them from the African or negro race. It is amazing that my articles on the Dravidians in Current Biology, and my paper on the Fulani in the PNAS, overturned the status quo.
The Dravidian article made it clear that geneticist were hiding data that proved that negroes in Africa and South India were genetically related as proven earlier by blood-type, culture and phenotypical features.
The article on the Fulani, proved that they were Africans, and not Black/Dark skinned whites. It also made it clear that M173, unified European and African populations that carried the R haplogroup. As a result, when Cruciani came out with his renaming of African R1, into V88, the presence of R1 could no longer be explained by a back migration. To remove the African carrier of M207, the carriers of M207 in Africa, were now classified as V45.
Everything would have been okay except for the fact that I published by article on the possible African origin of R1, due to the diversity of R haplogroups in Africa and the fact that M269 was carried by African populations in various parts of Africa that did not live in a continuous geographical area proved that the existence of R haplogroup in Africa, confirmed that the presence of R in Africa was not due to a back migration.
Establishment Geneticists can not erase the genomic evidence published in journals around the world. They can not deny the evidence for an African origin of R hg, because it does exist.
This is why Eurocentrists hate that my articles have been published on-line where everyone can read them. People like lioness and Montellano are useful to establishment researchers in maintaining the status quo. They are useful because real geneticist know that their phylogenies and dating for y-chromosome R and and mtDNA M, are wrong and that these haplogroups had to have been present in Africa before 50kya. As a result, if they attack my work in one of their journals, I would just write a response using their own papers to falsify their research.
That is why Montellano had to publish his paper at Academia edu, instead of a PLoS or PNAS, the genetic evidence proves his article lacks validity.
quote:Originally posted by Clyde Winters: [ The goal of lioness and Montellano is to use misdirection to maintain the myth Europeans--whites--carry a unique set of genes that separate them from the African or negro race. It is amazing that my articles on the Dravidians in Current Biology, and my paper on the Fulani in the PNAS, overturned the status quo.
LOL Look at Winters' s impact factor compared to real population geneticists at the bottom of the post. Winter's articles are not cited , by anyone except himself. Clyde please post a citation praising your work in any mainline genetic journal.
quote:
This is why Eurocentrists hate that my articles have been published on-line where everyone can read them. People like lioness and Montellano are useful to establishment researchers in maintaining the status quo. They are useful because real geneticist know that their phylogenies and dating for y-chromosome R and and mtDNA M, are wrong and that these haplogroups had to have been present in Africa before 50kya. As a result, if they attack my work in one of their journals, I would just write a response using their own papers to falsify their research.
That is why Montellano had to publish his paper at Academia edu, instead of a PLoS or PNAS, the genetic evidence proves his article lacks validity.
Most of the papers Winters is posting have been published in journals from India or Pakistan which are post-reviewed i.e. no one reviews them before publication (no, they don't check the references either). People reading the paper are supposed to write and critique it afterwards LOL. It's not worth the effort. Winter has yet to publish a peer-reviewed publication in a recognized mainline scientific journal. BTW letters to the editor and comments to other published articles are NOT peer-reviewed. People, who have published in peer-reviewed journals and are familiar with the process know the difference.
Winters:
quote:Firstly, I am proud to be an Afrocentric scholar. I have been one all my academic life.
LOL. You have to pay a fee to publish in just about all online journals. In most cases as much as $1400 or more .
Moreover, most papers in genetics are published in open access journals.
Not true, most of the influential and cited articles in genetics are published in mainline peer-reviewed journals for instance just a few of the ones I look at American Journal of Human Genetics, Annals of Human Genetics[,[Proceedings of the National Academy of Science, PNAS, European Journal of Genetics, Nature, Science, Nature Genetics,Nature Reviews-Genetics;[BMC Genetics ,BMC Genomics, Molecular Biology and Evolution, American Journal of Physical Evolution, etc. etc. this is just a tiny sample of the peer-reviewed journals on genetics- take a look at the list in "Entrez PubMed."
Winters:
quote:You are a joke. I have had articles published in BioEssay, PNAS, Science and etc., these are 'mainstream journals'. Where are your publications? You can find my population genetics articles here:
I have published in a couple of papers in Science and Current Anthropology as well as papers in Journal of the American Anthropological Society. Journal of the American Chemical Society, Ethnohistory, Medical Anthropology,Advances in Nursing Science,Yearbook of Physical Anthropology,Journal of Ethnomedicine, Ethnomedizin among others as well as peer reviewing dozens of papers for mainline peer-reviewed journals.
Analyzing Winters list of publications confirms my previous statement that he has not published a refereed article in a mainline refereed journal. There were a number of broken links in his list, but all of his contributions to mainline journals i.e. Bioassay, PNAS, Science, and PLoS Genetics were letters commenting on someone elses article. Letters and comments are NOT peer-reviewed, and are mostly ignored by the authors of the original papers. Winters is probably counting on the fact that most people are not familiar with academic publishing and can be bamboozled into thinking that his contributions to prestigious journals are peer-reviewed articles. Dont take my word, go to his list and verify that these contributions are letters.
I wrote the editors of the mainline journals where Winters published his comments, and as expected these things are NOT peer-reviewed before publication. For example from PLoS Genetics
quote: Subject: RE: editorial policy Date: Tue, 8 May 2007 07:58:12 -0700 From: "PLoS Genetics" <plosgenetics@plos.org> To: Dear Bernard, Thanks for your message good question. Reader Responses are intended to be more informal and to encourage community dialogue. As such, they do not undergo peer review by our editors or by external referees (whereas correspondence is treated differently and is peer reviewed).
Instead, Reader Responses are reviewed by staff (to check they are not obscene, abusive, defamatory, libelous, or in some other way illegal or discriminatory; otherwise, we will post them). I hope this helps.
Best wishes,
Andy Andy Collings Publications Manager, PLoS Genetics plosgenetics@plos.org / http://www.plosgenetics.org/ Email Alerts: " target="_blank">http://register.plos.org/
Most of the time authors dont even bother to reply to comments by Winters. When they do, the echo points Ive made repeatedly on this group.
Did the Dravidian Speakers Originate in Africa In a familiar pattern, the authors of the article Winters was commenting on point out that Winters is not familiar with genetics:
Chaubey, G. et al. 2007 Reply to Winters, BioAssays 29: 499.
quote:MtDNA-based genetic arguments provided by Dr. Winters in favor of gene flow from Africa to Dravidian-speaking Indians are, however, entirely erroneous. The author has been, unfortunately, confused by overlooking changes in mtDNA haplogroup (hg) nomenclature. Namely hg, M1 in Kivisild et al.(4) has been later changed to hg M3, in order to avoid parallel nomenclatures.(5) Furthermore, a recent dedicated paper on phylogeography of mtDNA hg M1(6) as well as an extensive comparative mapping of autosomal genetic markers among many Indian populations relative to global populations elsewhere, including Africans,(7) do not provide any clues for a putative recent gene flow, from Africa, to Dravidian-speaking populations in South Asia.
ResearchGate publishes a measure of what impact a particular scholars publications have that includes the number times articles are cited, seen and downloaded together with prestige of the journals they are published in equaling an impact factor
here are the scores for the four authors of the reply above, the author of the review of Winterss article starting this thread as well as Winters and my impact factor for comparison:
Name papers reads citations impact Toomas Kivisild 157 15K 9,444 1,133.89 R. Villems 179 15K 8,485 1,122.55 Mait Metzpalu 69 12K 2,870 665.13 G. Chaubey 53 6K 1,020 322.03 Klyosov, A. 96 3K 1,168 141.94 Bernard O de M 45 521 348 223.18 Clyde Winters 65 3K 92 56.22
Posts: 833 | From: Austin, TX | Registered: Jan 2007
| IP: Logged |
posted
A post-Review of a "paper" by Clyde Winters http://classic.rstb.royalsocietypublishing.org/content/367/1590/770.abstract/reply - content-block Haplogroup L3 (M,N) probably spread across Africa before the Out of Africa event Clyde Winters, Professor Uthman dan Fodio Institute Oppenheimer presents a convincing article supporting an Out of Africa (OoA) event [1]. At this time, beachcombing anatomically modern humans (amh) exploiting marine resources made their way to Australia by 42kya, after a bottleneck in India [1]. It was in Eurasia that, according to Oppenheimer, L3 (M,N) originated and haplogroups M and N returned to Africa via a back migration [1]. Soares et al. also argue that L3 (M,N and R) originated in Eurasia [2]. Oppenheimer dates L3 (M,N) to 83 kya [1], while Soares et al. [2] and Behar [3] argue that L3 originated around 70 kya and haplogroups M,N, and R appeared between 50-59 kya in Eurasia. The genetic phylogeny of L3 suggest that L3 (M,N) had probably spread across Africa before the Toba eruption and OoA event so it could not have originated in Eurasia. Haplogroup L3 (M,N) probably spread from East Africa to West Africa along with haplogroup LOd. The most recent common ancestor (TMRC) of AMH carrying LOd according to Gonder et al. dates to 106kya. A haplotype of LOd is AF-24, delineated by DdeI site 10394 and AluI site of np 10397. Haplogroup L3 (M,N) is characterized by the DdeI site np 10394 and AluI 10397. Chen et al. maintain that Haplotype AF-24 (DQ112852) is at the base of the M Haplogroup [4]. The only African mtDNA that has both of these sites is the Senegalese AF-24 associated with Haplogroup L3 [Chen]. The presence of AF-24 among the Senegalese, given the antiquity of this haplotype in relation to LOd, suggests an early expansion of L3 (M,N) from East Africa to West Africa before the OoA event suggested by Oppenheimer. Gonder et al. has dated L3 to 100kya [5]. Oppenheimer [1] and Behar et al [3] on the other hand have postulated an estimated time for TMRCA of L3 to around 70kya. The presence of L3 (M,N) in West Africa and haplotype AF- 24 suggest an ancient demic diffusion of L3 (M,N) to West Africa prior to 70kya, and support Soares et al.'s [2] and Gonder et al.'s [5] dating of L3 between 80-100kya. Anatomically modern humans arrived in Senegal during the Sangoan period. Sangoan artifacts spread from East Africa to West Africa between 100-80kya. In Senegal Sangoan material was discovered near Cap Manuel [6], Gambia River in Senegal [8,9]; and Cap Vert [7]. The distribution of the Sangoan culture supports the demic diffusion of L3 (M,N) into West Africa over 100kya. The spread of the Sangoan tool kit into the Senegambian region [6-9] would explain the presence of haplotype AF-24 among the Senegalese. This archaeological evidence adds additional corroboration to the West African genetic phylogenies which point to the probable expansion of L3(M,N) from East to West Africa prior to the OoA event indicated by Oppenheimer [1] 100 kya. References: 1. Oppenheimer S. 2012 Out-of-Africa, the peopling of continents and islands: tracing uniparental gene trees across the map. Phil. Trans. R. Soc. Lond. B, 367, 770-784. (doi: 10.1098/rstb.2011.0306) 2. Soares P., Ermini L., Thomson, N., Mormina M., Rito T., Rohl A., Salas A., Oppenheimer S., Macaulay V., Richards M.B. 2009 Correcting for purifying selection: an improved human mitochondrial molecular clock. Am. J. Hum. Genet., 84, 740-759. (doi:10.1016/j.ajhg.2009.05.001) 3. Oppenheimer S. 2009 The great arc of dispersal of modern humans: Africa to Australia. Quat. Int., 202, 2-13. (doi:10.1016/j.quaint.2008.05.015) 4. Chen Y-S., Olckers A., Schurr T.G., Kogelnik A.M., Huoponen K., Wallace D.C. 2000 mtDNA variation in the South African Kung and Khwe - and their genetic relationships to other African populations. Am. J. Hum. Genet., 66, 1362-1383. 5. Gonder M.K., Mortensen H.M., Reed F.A., de Sousa A., Tishkoff S.A. 2007 Whole mtDNA genome sequence analysis of ancient African lineages. Mol. Biol. Evol., 24, 757-768. (doi: 10.1093/molbev/msl209) 6. Giresse, P. 2008 Tropical and sub-Tropical West Africa - marine and continental changes during the Late Quaternary. Elsevier Science. 7. Phillipson, D.W. 2005 African Archaeology. Cambridge University Press, Cambridge. 8. Davies, O. 1967 West Africa before the Europeans. Methuen, London. 9. Wai-Ogusu, A. 1973 Was there a Sangoan industry in West Africa. West Afr. J. Arcah., 3, 191-96.
%%%%%%%%%%%% Response to Winters (2012) 'Haplogroup L3 (M,N) probably spread across Africa before the Out of Africa event' Stephen Oppenheimer School of Anthropology and Museum Ethnography, Institute of Cognitive and Evolutionary Anthropology, Winters' response to my recent review in Phil. Trans. R. Soc. B [1] argues that the two mtDNA super-haplogroups M and N (which account for nearly all non-Africans and derive from African haplogroup L3) both originated within Africa, before the anatomically modern human (AMH) 'Out of Africa' (OOA) exit, and further that African L3 dates to 100kya (the latter date being based on a misreading of the cited sources). His argument rests mainly on the acknowledged phylogenetic ambiguity of a single Senegalese sample in a paper published in 2000 before complete sequencing was routinely used to resolve such ambiguities. Local African origins of both N & M have been argued before on the basis of the presence of M sub-group M1 and certain derivative N sub- groups in East Africa; and this evidence of local backflow has already been clarified in the modern literature - as discussed in my review [1]. While the said N sub-groups are clearly derived from N outside Africa and are much younger than any postulated exit date, African sub-group M1 is, perhaps, more interesting having a rather greater age than N in Africa, though still considerably younger than any estimate of the OOA event [1] and M1 is present, though rare [2], outside Africa, consistent with an ex- African origin. Genetic phylogeographic arguments contribute greatly to the topographical inference of the sequence of migrations and, independently of any date estimates, argue strongly for M & N originating outside Africa from a single L3 exit [1]. MtDNA dates are presently more reliable than for any other genetic locus but, as discussed exhaustively in my review [1], their high calculated variance still gives wide error bars and the opportunity for much fruitless argument on precise dates of AMH exit either side of the Toba volcanic event, which could best be resolved by diagnostic, accurately dated modern human fossils (rather than stone tools) at key points outside Africa. For the dates suggested by Winters, such early AMH fossils are currently lacking. As for the chosen genetic dates he cites for his own preferred chronology, several of which are misquoted and/or misunderstood, those from the older literature have been superseded by more recent, more resolved, recalibrated, comprehensive, accurate and precise studies on L3 within and outside Africa, including M and N [3, 4], including one major source Winters suggests himself [5]. Finally, there are is a string of misunderstandings mistakes and misquotes in Winters' letter listed below, which detract from, rather than supporting, his overall argument: Para 1, sentence 2: 42 kya refers presumably to the earliest carbon dates in Timor, not those much earlier luminescence dates given, in my review [1], for Australia. Para 2, sentence 1: 'Oppenheimer dates L3 (M,N) to 83 kya' Comment: I did not do so in this review [1]. As explained in the text I chose throughout to cite lineage ages from the key mtDNA recalibration paper of Soares et al. (2009) [5]. This was both for consistency, and to use the latest, most comprehensive and, hopefully, least inaccurate method, rather than pick and choose results from older, phylogenetically less-resolved publications, which might perhaps have suited my own preference for a pre-Toba exit better. Para 3 (whole para): Comment: No published evidence/reference given for these assertions. Para 4, sentence 1: 'The most recent common ancestor (TMRC) of AMH carrying LOd according to Gonder et al. dates to 106kya.'. Comment: 'LOd' is not 'TMRC of AMH' (nor is L0d). Para 4, sentence 1: 'A haplotype of LOd is AF-24...' Comment: No it is not - on the evidence given in the citation. Gonder et al. (2007) [6] do not even mention haplotype AF-24 as such. The claim, if it were true, would simply reinforce the impression of ambiguity. AF-24 is however mentioned by Winters' other cited reference (published in 2000 and not based on complete sequences): Chen et al. (2000) [7] shown as belonging to L3a; but those authors acknowledge this particular phylogenetic assignment to be poorly resolved: 'Haplotype AF24, which is aligned with Asian macrohaplogroup M, is indicated by a double section symbol...subclusters AF19-AF21/AF24 and AF80-AF84 were not resolved at bootstrap values >50%...' [7] They further emphasize the ambiguity of its phylogenetic/geographic assignment: '...it is also possible that this particular haplotype [AF-24] is present in Africa because of back-migration [of M] from Asia.'... and: 'Alternatively, AF24 may have been introduced from Asia into Africa more recently.' [7] Para 4, sentence 3: Winters continues to mis-cite Chen et al. (2000) [7]: 'Chen et al. maintain that Haplotype AF-24 (DQ112852) is at the base of the M Haplogroup [4].', Comment: Where? - unless it is in their reference to back-migration (above). Para 5 appears to be further argument based on phylogenetic ambiguity. Para 6, sentence 1: 'Gonder et al. has dated L3 to 100kya (5).' Comment: No, they do not, according to Gonder's Table 2 [6]. Para 6, sentence 3: 'The presence of L3 (M,N) in West Africa and haplotype AF- 24 suggest an ancient demic diffusion of L3 (M,N) to West Africa prior to 70kya, and support Soares et al.'s (2) and Gonder et al.'s (5) dating of L3 between 80-100kya.' Comment: The above inference, based on a single poorly-resolved haplotype, is unsound and Soares et al. (2009) [5] are mis-cited as far as the date is concerned. Paras 7 and 8: Varied, inadequately-cited references to the presence of the 'Sangoan tool kit' in West Africa are used by Winters to infer the movement and spread of 'L3 (M,N)' in West Africa. Comment: This is an unwarranted inference using as it does, hypothetical links (for which no evidence is given) between an Early Stone Age cultural phase and an ambiguous single modern genetic haplotype (AF-24). While dated archaeological evidence of human presence can, occasionally, be used to infer first-ever human arrival in a previously uninhabited region e.g. Polynesia or the Canary Islands and this kind of evidence be used to cross -check calibration of the mtDNA clock on unique and specific local founding lineages in those places [5], the sort of "stones and genes" type of inferences Winters makes for the spread of 'L3 (M,N)' in West Africa are completely unwarranted. References 1. Oppenheimer S. 2012 Out-of-Africa, the peopling of continents and islands: tracing uniparental gene trees across the map. Phil. Trans. R. Soc. Lond. B, 367, 770-784. (doi: 10.1098/rstb.2011.0306) 2. Gunnarsdottir E.D., Nandineni M.R., Li M., Myles S., Gil D., Pakendorf B., Stoneking M. (2011) Larger mitochondrial DNA than Y- chromosome differences between matrilocal and patrilocal groups from Sumatra. Nature Communications, 2, 228. (doi: 10.1038/ncomms1235) 3. Soares P., Alshamali F., Pereira J.B., Fernandes, V., N.M. Silva, Afonso C., Costa M.D., Musilova E., Macaulay V., Richards M.B., Cerny V., Pereira Luisa. (2012) The expansion of mtDNA haplogroup L3 within and out of Africa. Mol. Biol. Evol., 29, 915-927. (doi: 10.1093/molbev/msr245) 4. Fernandes, V., Alshamali F., Alves M., Costa M.D., Pereira J.B., N.M. Silva, Cherni L., Harich N., Cerny V., Soares P. Richards M.B., Pereira Luisa. (2012) The Arabian cradle: Mitochondrial relicts of the first steps along the Southern Route out of Africa. Am. J. Hum. Genet., 90, 347-355. (doi: 10.1016/j.ajhg.2011.12.010) 5. Soares P., Ermini L., Thomson, N., Mormina M., Rito T., Rohl A., Salas A., Oppenheimer S., Macaulay V., Richards M.B. 2009 Correcting for purifying selection: an improved human mitochondrial molecular clock. Am. J. Hum. Genet., 84, 740-759. (doi:10.1016/j.ajhg.2009.05.001) 6. Gonder M.K., Mortensen H.M., Reed F.A., de Sousa A., Tishkoff S.A. 2007 Whole mtDNA genome sequence analysis of ancient African lineages. Mol. Biol. Evol., 24, 757-768. (doi: 10.1093/molbev/msl209) 7. Chen Y-S., Olckers A., Schurr T.G., Kogelnik A.M., Huoponen K., Wallace D.C. 2000 mtDNA variation in the South African Kung and Khwe - and their genetic relationships to other African populations. Am. J. Hum. Genet., 66, 1362-1383.
Posts: 833 | From: Austin, TX | Registered: Jan 2007
| IP: Logged |
quote:Originally posted by Quetzalcoatl: A post-Review of a "paper" by Clyde Winters http://classic.rstb.royalsocietypublishing.org/content/367/1590/770.abstract/reply - content-block Haplogroup L3 (M,N) probably spread across Africa before the Out of Africa event Clyde Winters, Professor Uthman dan Fodio Institute Oppenheimer presents a convincing article supporting an Out of Africa (OoA) event [1]. At this time, beachcombing anatomically modern humans (amh) exploiting marine resources made their way to Australia by 42kya, after a bottleneck in India [1]. It was in Eurasia that, according to Oppenheimer, L3 (M,N) originated and haplogroups M and N returned to Africa via a back migration [1]. Soares et al. also argue that L3 (M,N and R) originated in Eurasia [2]. Oppenheimer dates L3 (M,N) to 83 kya [1], while Soares et al. [2] and Behar [3] argue that L3 originated around 70 kya and haplogroups M,N, and R appeared between 50-59 kya in Eurasia. The genetic phylogeny of L3 suggest that L3 (M,N) had probably spread across Africa before the Toba eruption and OoA event so it could not have originated in Eurasia. Haplogroup L3 (M,N) probably spread from East Africa to West Africa along with haplogroup LOd. The most recent common ancestor (TMRC) of AMH carrying LOd according to Gonder et al. dates to 106kya. A haplotype of LOd is AF-24, delineated by DdeI site 10394 and AluI site of np 10397. Haplogroup L3 (M,N) is characterized by the DdeI site np 10394 and AluI 10397. Chen et al. maintain that Haplotype AF-24 (DQ112852) is at the base of the M Haplogroup [4]. The only African mtDNA that has both of these sites is the Senegalese AF-24 associated with Haplogroup L3 [Chen]. The presence of AF-24 among the Senegalese, given the antiquity of this haplotype in relation to LOd, suggests an early expansion of L3 (M,N) from East Africa to West Africa before the OoA event suggested by Oppenheimer. Gonder et al. has dated L3 to 100kya [5]. Oppenheimer [1] and Behar et al [3] on the other hand have postulated an estimated time for TMRCA of L3 to around 70kya. The presence of L3 (M,N) in West Africa and haplotype AF- 24 suggest an ancient demic diffusion of L3 (M,N) to West Africa prior to 70kya, and support Soares et al.'s [2] and Gonder et al.'s [5] dating of L3 between 80-100kya. Anatomically modern humans arrived in Senegal during the Sangoan period. Sangoan artifacts spread from East Africa to West Africa between 100-80kya. In Senegal Sangoan material was discovered near Cap Manuel [6], Gambia River in Senegal [8,9]; and Cap Vert [7]. The distribution of the Sangoan culture supports the demic diffusion of L3 (M,N) into West Africa over 100kya. The spread of the Sangoan tool kit into the Senegambian region [6-9] would explain the presence of haplotype AF-24 among the Senegalese. This archaeological evidence adds additional corroboration to the West African genetic phylogenies which point to the probable expansion of L3(M,N) from East to West Africa prior to the OoA event indicated by Oppenheimer [1] 100 kya. References: 1. Oppenheimer S. 2012 Out-of-Africa, the peopling of continents and islands: tracing uniparental gene trees across the map. Phil. Trans. R. Soc. Lond. B, 367, 770-784. (doi: 10.1098/rstb.2011.0306) 2. Soares P., Ermini L., Thomson, N., Mormina M., Rito T., Rohl A., Salas A., Oppenheimer S., Macaulay V., Richards M.B. 2009 Correcting for purifying selection: an improved human mitochondrial molecular clock. Am. J. Hum. Genet., 84, 740-759. (doi:10.1016/j.ajhg.2009.05.001) 3. Oppenheimer S. 2009 The great arc of dispersal of modern humans: Africa to Australia. Quat. Int., 202, 2-13. (doi:10.1016/j.quaint.2008.05.015) 4. Chen Y-S., Olckers A., Schurr T.G., Kogelnik A.M., Huoponen K., Wallace D.C. 2000 mtDNA variation in the South African Kung and Khwe - and their genetic relationships to other African populations. Am. J. Hum. Genet., 66, 1362-1383. 5. Gonder M.K., Mortensen H.M., Reed F.A., de Sousa A., Tishkoff S.A. 2007 Whole mtDNA genome sequence analysis of ancient African lineages. Mol. Biol. Evol., 24, 757-768. (doi: 10.1093/molbev/msl209) 6. Giresse, P. 2008 Tropical and sub-Tropical West Africa - marine and continental changes during the Late Quaternary. Elsevier Science. 7. Phillipson, D.W. 2005 African Archaeology. Cambridge University Press, Cambridge. 8. Davies, O. 1967 West Africa before the Europeans. Methuen, London. 9. Wai-Ogusu, A. 1973 Was there a Sangoan industry in West Africa. West Afr. J. Arcah., 3, 191-96.
%%%%%%%%%%%% Response to Winters (2012) 'Haplogroup L3 (M,N) probably spread across Africa before the Out of Africa event' Stephen Oppenheimer School of Anthropology and Museum Ethnography, Institute of Cognitive and Evolutionary Anthropology, Winters' response to my recent review in Phil. Trans. R. Soc. B [1] argues that the two mtDNA super-haplogroups M and N (which account for nearly all non-Africans and derive from African haplogroup L3) both originated within Africa, before the anatomically modern human (AMH) 'Out of Africa' (OOA) exit, and further that African L3 dates to 100kya (the latter date being based on a misreading of the cited sources). His argument rests mainly on the acknowledged phylogenetic ambiguity of a single Senegalese sample in a paper published in 2000 before complete sequencing was routinely used to resolve such ambiguities. Local African origins of both N & M have been argued before on the basis of the presence of M sub-group M1 and certain derivative N sub- groups in East Africa; and this evidence of local backflow has already been clarified in the modern literature - as discussed in my review [1]. While the said N sub-groups are clearly derived from N outside Africa and are much younger than any postulated exit date, African sub-group M1 is, perhaps, more interesting having a rather greater age than N in Africa, though still considerably younger than any estimate of the OOA event [1] and M1 is present, though rare [2], outside Africa, consistent with an ex- African origin. Genetic phylogeographic arguments contribute greatly to the topographical inference of the sequence of migrations and, independently of any date estimates, argue strongly for M & N originating outside Africa from a single L3 exit [1]. MtDNA dates are presently more reliable than for any other genetic locus but, as discussed exhaustively in my review [1], their high calculated variance still gives wide error bars and the opportunity for much fruitless argument on precise dates of AMH exit either side of the Toba volcanic event, which could best be resolved by diagnostic, accurately dated modern human fossils (rather than stone tools) at key points outside Africa. For the dates suggested by Winters, such early AMH fossils are currently lacking. As for the chosen genetic dates he cites for his own preferred chronology, several of which are misquoted and/or misunderstood, those from the older literature have been superseded by more recent, more resolved, recalibrated, comprehensive, accurate and precise studies on L3 within and outside Africa, including M and N [3, 4], including one major source Winters suggests himself [5]. Finally, there are is a string of misunderstandings mistakes and misquotes in Winters' letter listed below, which detract from, rather than supporting, his overall argument: Para 1, sentence 2: 42 kya refers presumably to the earliest carbon dates in Timor, not those much earlier luminescence dates given, in my review [1], for Australia. Para 2, sentence 1: 'Oppenheimer dates L3 (M,N) to 83 kya' Comment: I did not do so in this review [1]. As explained in the text I chose throughout to cite lineage ages from the key mtDNA recalibration paper of Soares et al. (2009) [5]. This was both for consistency, and to use the latest, most comprehensive and, hopefully, least inaccurate method, rather than pick and choose results from older, phylogenetically less-resolved publications, which might perhaps have suited my own preference for a pre-Toba exit better. Para 3 (whole para): Comment: No published evidence/reference given for these assertions. Para 4, sentence 1: 'The most recent common ancestor (TMRC) of AMH carrying LOd according to Gonder et al. dates to 106kya.'. Comment: 'LOd' is not 'TMRC of AMH' (nor is L0d). Para 4, sentence 1: 'A haplotype of LOd is AF-24...' Comment: No it is not - on the evidence given in the citation. Gonder et al. (2007) [6] do not even mention haplotype AF-24 as such. The claim, if it were true, would simply reinforce the impression of ambiguity. AF-24 is however mentioned by Winters' other cited reference (published in 2000 and not based on complete sequences): Chen et al. (2000) [7] shown as belonging to L3a; but those authors acknowledge this particular phylogenetic assignment to be poorly resolved: 'Haplotype AF24, which is aligned with Asian macrohaplogroup M, is indicated by a double section symbol...subclusters AF19-AF21/AF24 and AF80-AF84 were not resolved at bootstrap values >50%...' [7] They further emphasize the ambiguity of its phylogenetic/geographic assignment: '...it is also possible that this particular haplotype [AF-24] is present in Africa because of back-migration [of M] from Asia.'... and: 'Alternatively, AF24 may have been introduced from Asia into Africa more recently.' [7] Para 4, sentence 3: Winters continues to mis-cite Chen et al. (2000) [7]: 'Chen et al. maintain that Haplotype AF-24 (DQ112852) is at the base of the M Haplogroup [4].', Comment: Where? - unless it is in their reference to back-migration (above). Para 5 appears to be further argument based on phylogenetic ambiguity. Para 6, sentence 1: 'Gonder et al. has dated L3 to 100kya (5).' Comment: No, they do not, according to Gonder's Table 2 [6]. Para 6, sentence 3: 'The presence of L3 (M,N) in West Africa and haplotype AF- 24 suggest an ancient demic diffusion of L3 (M,N) to West Africa prior to 70kya, and support Soares et al.'s (2) and Gonder et al.'s (5) dating of L3 between 80-100kya.' Comment: The above inference, based on a single poorly-resolved haplotype, is unsound and Soares et al. (2009) [5] are mis-cited as far as the date is concerned. Paras 7 and 8: Varied, inadequately-cited references to the presence of the 'Sangoan tool kit' in West Africa are used by Winters to infer the movement and spread of 'L3 (M,N)' in West Africa. Comment: This is an unwarranted inference using as it does, hypothetical links (for which no evidence is given) between an Early Stone Age cultural phase and an ambiguous single modern genetic haplotype (AF-24). While dated archaeological evidence of human presence can, occasionally, be used to infer first-ever human arrival in a previously uninhabited region e.g. Polynesia or the Canary Islands and this kind of evidence be used to cross -check calibration of the mtDNA clock on unique and specific local founding lineages in those places [5], the sort of "stones and genes" type of inferences Winters makes for the spread of 'L3 (M,N)' in West Africa are completely unwarranted. References 1. Oppenheimer S. 2012 Out-of-Africa, the peopling of continents and islands: tracing uniparental gene trees across the map. Phil. Trans. R. Soc. Lond. B, 367, 770-784. (doi: 10.1098/rstb.2011.0306) 2. Gunnarsdottir E.D., Nandineni M.R., Li M., Myles S., Gil D., Pakendorf B., Stoneking M. (2011) Larger mitochondrial DNA than Y- chromosome differences between matrilocal and patrilocal groups from Sumatra. Nature Communications, 2, 228. (doi: 10.1038/ncomms1235) 3. Soares P., Alshamali F., Pereira J.B., Fernandes, V., N.M. Silva, Afonso C., Costa M.D., Musilova E., Macaulay V., Richards M.B., Cerny V., Pereira Luisa. (2012) The expansion of mtDNA haplogroup L3 within and out of Africa. Mol. Biol. Evol., 29, 915-927. (doi: 10.1093/molbev/msr245) 4. Fernandes, V., Alshamali F., Alves M., Costa M.D., Pereira J.B., N.M. Silva, Cherni L., Harich N., Cerny V., Soares P. Richards M.B., Pereira Luisa. (2012) The Arabian cradle: Mitochondrial relicts of the first steps along the Southern Route out of Africa. Am. J. Hum. Genet., 90, 347-355. (doi: 10.1016/j.ajhg.2011.12.010) 5. Soares P., Ermini L., Thomson, N., Mormina M., Rito T., Rohl A., Salas A., Oppenheimer S., Macaulay V., Richards M.B. 2009 Correcting for purifying selection: an improved human mitochondrial molecular clock. Am. J. Hum. Genet., 84, 740-759. (doi:10.1016/j.ajhg.2009.05.001) 6. Gonder M.K., Mortensen H.M., Reed F.A., de Sousa A., Tishkoff S.A. 2007 Whole mtDNA genome sequence analysis of ancient African lineages. Mol. Biol. Evol., 24, 757-768. (doi: 10.1093/molbev/msl209) 7. Chen Y-S., Olckers A., Schurr T.G., Kogelnik A.M., Huoponen K., Wallace D.C. 2000 mtDNA variation in the South African Kung and Khwe - and their genetic relationships to other African populations. Am. J. Hum. Genet., 66, 1362-1383.
This is my response to Oppenheimer
MONDAY, JUNE 11, 2012 Response by Oppenheimer to Winters'(2012) Haplogroup L3(M,N) probably spread across Africa before the Out of Africa Event Dr. Oppenheimer (2012b) implies that L3(M,N) originated in Asia. This is false. We know that L3 originated long before the OoA event. He does not present any evidence falsifying my conclusion. His entire argument is that M1 is rare in Asia.
Haplotypes with HVSI transitions defining 16129-16223-16249-16278-16311-16362; and 16129-16223-16234-16249-16211-16362 have been found in Thailand and among the Han Chinese (Fucharoen et al., 2001; Yao et al., 2002) and these were originally thought to be members of Haplogroup M1. However, on the basis of currently available FGS sequences, carriers of these markers have been found to be in the D4a branch of Haplogroup D, the most widespread branch of M 1 in East Asia (Fucharoen et al., 2001; Yao et al., 2002). The transitions 16129, 16189, 16249 and 16311 are known to be recurrent in various branches of Haplogroup M, especially M1 and D4.
Dr. Oppenheimer (2012b)claims that there are a string of mistakes and misquotes in my response to his article which are not substantiated by the literature. For example, Dr. Oppenheimer claims that LOd is not the TMRC for AMH. This is false. I claim that LOd is older than L3, this is not contradicted by Soares.
Atkinson et al (2009) makes it clear that L3 is the youngest African haplogroup and LO is the oldest. As a result, when Dr. Oppenheimer claims that LOd is not the TMRC of AMH, he is false. LO is the oldest haplogroup, since LOd is dated to 106kya and one of the LO clades it. Atkinson et al (2009) observed that Haplogroups L0 and L1 (figure 2b,c, respectively) show slow constant growth over the last 100200 kyr (TMRCAs: L0, 124172 kyr ago; L1, 87139 kyr ago; L0 and L1 combined, 156213 kyr ago; 95% HPDs). This makes it clear that haplogroup LO is the oldest mtDNA haplogroup in Africa.
Dr. Oppenheimer also claims that haplotype AF-24 is poorly resolved. This is false, Chen et al make it clear that The samples included complete haplotypes of 62 Senegalese (AF01AF24, AF26AF36, AF45AF59, AF64AF65, and AF70AF79). As a result, how can he make the claim AF-24 is poorly resolved when Chen et al (2002) make it clear that he used complete haplotypes of 62 Senegalese samples that include AF-24.
Chen et al makes it clear that AF-24 could be of either Asian or African originSimilarly, L3a was found to have a close affinity to haplotype AF24, a mtDNA that has the DdeI np-10394 and AluI np-10397 site gains characteristic of Asian macrohaplogroup M (figs.(figs.22 andand3).3). Therefore, it is possible that subhaplogroup L3a was the progenitor of Asian mtDNAs belonging to M. Although the age of subhaplogroup L3a is somewhat less than our estimate for the age of Asian haplogroup M (Torroni et al. 1994b; Chen et al. 1995), the differences could be due to the limited number of L3a mtDNAs in our African sample. Alternatively, AF24 may have been introduced from Asia into Africa more recently. The fact that Atkinson et al (2009) makes it clear that AF-24 is a haplotype of LO, make it unlikely that AF-24 originated in Asia, since it was already in existence prior to the OoA event.
Finally, Oppenheimer claims that you can not infer population movements relating to the expansion of the ancient tool kits. This is a false statement since the demic expansion of LO(d) and L3 from East Africa to West Africa is cross referenced with specific founding lineage which is assumed to have originated in the East. This assumption is just as valid as Oppenheimers view relating to the Tonga events impact on the OoA.
In summary, it is obvious that Dr. Oppenheimer has little knowledge of the expansion of haplogroups in Africa. I am surprised the he didnt know that the GenBank Accession number for Haplotype AF-24 is DQ112852, this suggest that he is not keeping up with the literature. Moreover, the earliest examples of L3(N) come from Iberia, not East Asia. Since this area was first occupied by Neanderthals until the expansion of the Aurignacian culture which had to have crossed the Straits of Gibraltar from Africa (Winters,2012). No where in Dr. Oppenhiemers response dose he present textual evidence supporting his conclusions. He only provides his opinionsnot evidence.
Chen Y-S., Olckers A., Schurr T.G., Kogelnik A.M., Huoponen K., Wallace D.C. 2000 mtDNA variation in the South African Kung and Khwe - and their genetic relationships to other African populations. Am. J. Hum. Genet., 66, 1362-1383
Fucharoen, G., S. Fucharoen and S. Horai, 2001. Mitochondrial DNA polymorphism in Thailand. J.Hum. Genet., 46: 115-125.
Gonder M.K., Mortensen H.M., Reed F.A., de Sousa A., Tishkoff S.A. 2007 Whole mtDNA genome sequence analysis of ancient African lineages. Mol. Biol. Evol., 24, 757-768. (doi: 10.1093/molbev/msl209).
Oppenheimer S. 2012 Out-of-Africa, the peopling of continents and islands: tracing uniparental gene trees across the map. Phil. Trans. R. Soc. Lond. B, 367, 770-784. (doi: 10.1098/rstb.2011.0306
Soares P., Ermini L., Thomson, N., Mormina M., Rito T., Rohl A., Salas A., Oppenheimer S., Macaulay V., Richards M.B. 2009 Correcting for purifying selection: an improved human mitochondrial molecular clock. Am. J. Hum. Genet., 84, 740-759. (doi:10.1016/j.ajhg.2009.05.001)
Yao, Y.G., Q.P. Kong, H.J. Bandelt, T. Kivisild and Y.P.Zhang, 2002. Phylogeographic differentiation of mitochondrial DNA in Han chinese. Am. J. Hum.Genet., 70: 635-651.
Posts: 13012 | From: Chicago | Registered: Jan 2006
| IP: Logged |
Are these the papers you are referring at, when you spoke of the M1-M3 transposition?
4. Kivisild T, Kaldma K, Metspalu M, Parik J, Papiha S, Villems R. 1999. The place of the Indian mtDNA variants in the global network of maternal lineages and the peopling of the Old World. In Deka R, Papiha SS. eds Genomic Diversity Kluwer Academic/Plenum Publishers New York p 135152.
5. Quintana-Murci L, Semino O, Bandelt H-J., Passarino G, McElreavey K, Santachiara-Benerecetti AS. 1999. Genetic evidence of an early exit of Homo sapiens sapiens from Africa through eastern Africa. Nat Genet 23: 437 441.
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
quote:Originally posted by Clyde Winters: [ The goal of lioness and Montellano is to use misdirection to maintain the myth Europeans--whites--carry a unique set of genes that separate them from the African or negro race. It is amazing that my articles on the Dravidians in Current Biology, and my paper on the Fulani in the PNAS, overturned the status quo.
LOL Look at Winters' s impact factor compared to real population geneticists at the bottom of the post. Winter's articles are not cited , by anyone except himself. Clyde please post a citation praising your work in any mainline genetic journal.
quote:
This is why Eurocentrists hate that my articles have been published on-line where everyone can read them. People like lioness and Montellano are useful to establishment researchers in maintaining the status quo. They are useful because real geneticist know that their phylogenies and dating for y-chromosome R and and mtDNA M, are wrong and that these haplogroups had to have been present in Africa before 50kya. As a result, if they attack my work in one of their journals, I would just write a response using their own papers to falsify their research.
That is why Montellano had to publish his paper at Academia edu, instead of a PLoS or PNAS, the genetic evidence proves his article lacks validity.
Most of the papers Winters is posting have been published in journals from India or Pakistan which are post-reviewed i.e. no one reviews them before publication (no, they don't check the references either). People reading the paper are supposed to write and critique it afterwards LOL. It's not worth the effort. Winter has yet to publish a peer-reviewed publication in a recognized mainline scientific journal. BTW letters to the editor and comments to other published articles are NOT peer-reviewed. People, who have published in peer-reviewed journals and are familiar with the process know the difference.
Winters:
quote:Firstly, I am proud to be an Afrocentric scholar. I have been one all my academic life.
LOL. You have to pay a fee to publish in just about all online journals. In most cases as much as $1400 or more .
Moreover, most papers in genetics are published in open access journals.
Not true, most of the influential and cited articles in genetics are published in mainline peer-reviewed journals for instance just a few of the ones I look at American Journal of Human Genetics, Annals of Human Genetics[,[Proceedings of the National Academy of Science, PNAS, European Journal of Genetics, Nature, Science, Nature Genetics,Nature Reviews-Genetics;[BMC Genetics ,BMC Genomics, Molecular Biology and Evolution, American Journal of Physical Evolution, etc. etc. this is just a tiny sample of the peer-reviewed journals on genetics- take a look at the list in "Entrez PubMed."
Winters:
quote:You are a joke. I have had articles published in BioEssay, PNAS, Science and etc., these are 'mainstream journals'. Where are your publications? You can find my population genetics articles here:
I have published in a couple of papers in Science and Current Anthropology as well as papers in Journal of the American Anthropological Society. Journal of the American Chemical Society, Ethnohistory, Medical Anthropology,Advances in Nursing Science,Yearbook of Physical Anthropology,Journal of Ethnomedicine, Ethnomedizin among others as well as peer reviewing dozens of papers for mainline peer-reviewed journals.
Analyzing Winters list of publications confirms my previous statement that he has not published a refereed article in a mainline refereed journal. There were a number of broken links in his list, but all of his contributions to mainline journals i.e. Bioassay, PNAS, Science, and PLoS Genetics were letters commenting on someone elses article. Letters and comments are NOT peer-reviewed, and are mostly ignored by the authors of the original papers. Winters is probably counting on the fact that most people are not familiar with academic publishing and can be bamboozled into thinking that his contributions to prestigious journals are peer-reviewed articles. Dont take my word, go to his list and verify that these contributions are letters.
I wrote the editors of the mainline journals where Winters published his comments, and as expected these things are NOT peer-reviewed before publication. For example from PLoS Genetics
quote: Subject: RE: editorial policy Date: Tue, 8 May 2007 07:58:12 -0700 From: "PLoS Genetics" <plosgenetics@plos.org> To: Dear Bernard, Thanks for your message good question. Reader Responses are intended to be more informal and to encourage community dialogue. As such, they do not undergo peer review by our editors or by external referees (whereas correspondence is treated differently and is peer reviewed).
Instead, Reader Responses are reviewed by staff (to check they are not obscene, abusive, defamatory, libelous, or in some other way illegal or discriminatory; otherwise, we will post them). I hope this helps.
Best wishes,
Andy Andy Collings Publications Manager, PLoS Genetics plosgenetics@plos.org / http://www.plosgenetics.org/ Email Alerts: " target="_blank">http://register.plos.org/
Most of the time authors dont even bother to reply to comments by Winters. When they do, the echo points Ive made repeatedly on this group.
Did the Dravidian Speakers Originate in Africa In a familiar pattern, the authors of the article Winters was commenting on point out that Winters is not familiar with genetics:
Chaubey, G. et al. 2007 Reply to Winters, BioAssays 29: 499.
quote:MtDNA-based genetic arguments provided by Dr. Winters in favor of gene flow from Africa to Dravidian-speaking Indians are, however, entirely erroneous. The author has been, unfortunately, confused by overlooking changes in mtDNA haplogroup (hg) nomenclature. Namely hg, M1 in Kivisild et al.(4) has been later changed to hg M3, in order to avoid parallel nomenclatures.(5) Furthermore, a recent dedicated paper on phylogeography of mtDNA hg M1(6) as well as an extensive comparative mapping of autosomal genetic markers among many Indian populations relative to global populations elsewhere, including Africans,(7) do not provide any clues for a putative recent gene flow, from Africa, to Dravidian-speaking populations in South Asia.
ResearchGate publishes a measure of what impact a particular scholars publications have that includes the number times articles are cited, seen and downloaded together with prestige of the journals they are published in equaling an impact factor
here are the scores for the four authors of the reply above, the author of the review of Winterss article starting this thread as well as Winters and my impact factor for comparison:
Name papers reads citations impact Toomas Kivisild 157 15K 9,444 1,133.89 R. Villems 179 15K 8,485 1,122.55 Mait Metzpalu 69 12K 2,870 665.13 G. Chaubey 53 6K 1,020 322.03 Klyosov, A. 96 3K 1,168 141.94 Bernard O de M 45 521 348 223.18 Clyde Winters 65 3K 92 56.22
You're just jealous that I have peer reviewed articles published eventhough I am an Afrocentric researcher.It is great news that Professional Scientists in India and Pakistan have the nerve to publish their own peer reviewed journals.
For years we had journals that published Afrocentric research including the Journal of Negro History, Negro History Bulletin, Black World and later Journal of African Civilization. Sadly, today we don't have any professional or amateur historians and anthropologists writing serious research in Afrocentric studies. In fact, active Afrocentric research is only taking place at this site. We have a lot of speakers talking about Afrocantric research dating back to the 1970's, but no real research into Afrocentric history and anthropology themes.
Today, I am the only one publishing Afrocentric studies in research journals, Mike has created a great Website on Black and African History, while Ironlion publishes Afrocentric articles. We need more people with the nerve to publish more Afrocentric studies.
Eventhough my papers are suppose to have a low impact, it is funny that since my publications appeared, the phylogeny charts have been changed to make it appear that M207 and V45 are now listed as different haplogroups ; and people now are aware of the fact that Dravidians do in fact carry M1.
I will make a prediction that soon researchers will change the name of M269 in Africa.
Posts: 13012 | From: Chicago | Registered: Jan 2006
| IP: Logged |
posted
Quetzalcoatl, is like. It's only peer reviewed if (whites) the western world has reviewed and consigned it.
It's somewhat the mindset of a colonist.
What Quetzalcoatl is saying is that western academia is superior, therefore credible.
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
quote:Originally posted by Troll Patrol # Ish Gebor: @ Quetzalcoatl,
Are these the papers you are referring at, when you spoke of the M1-M3 transposition?
4. Kivisild T, Kaldma K, Metspalu M, Parik J, Papiha S, Villems R. 1999. The place of the Indian mtDNA variants in the global network of maternal lineages and the peopling of the Old World. In Deka R, Papiha SS. eds Genomic Diversity Kluwer Academic/Plenum Publishers New York p 135152.
5. Quintana-Murci L, Semino O, Bandelt H-J., Passarino G, McElreavey K, Santachiara-Benerecetti AS. 1999. Genetic evidence of an early exit of Homo sapiens sapiens from Africa through eastern Africa. Nat Genet 23: 437 441.
Yes, from my paper in Academia.edu
quote: . Kivisild (1999) and Quintana-Murci (1999) identified and named different haplotypes, one Indian and one African, as M1. This happens often and the usual procedure is for one of the authors to change the nomenclature. The final arbiter is www.phylotree.org that publishes an updated mtDNA tree. In this case, Quintana-Murcis (1999) identification of M1 was retained with haplotype markers 16129 16189 16249 16311. Kivisilds M1 had 16126 as the marker and this haplogroups name was changed to M3. The individuals in Kivisild (1999) labeled M1 did not disappear their designation changed to M3. Kivisilds compliance with the revised nomenclature is seen in Bamshad (2001). Numerous papers including some cited by Winters make it clear that there is no M1 in India although M3 with the 16126 marker (1999s M1) is definitely found (Bamshad 2001; Maca-Meyer 2001; Kivisild 2003- Dravidians; Metspalu 2004; Rajkumar 2005; Thangaraj 2006) Chandrasekar 2009; Shah 2012). In 2007, Winters (2007) repeated this distortion in a comment to a paper by Chaubey (2006). Chaubey, Metspalu, Kivisild, and Villems (2007) replied and clearly pointed out Winters misreporting of their paper. In 2008, Kivisild and Villems pointed this error to Winters by e-mail. Yet, he continues to use this argument and citation. This is academic dishonesty.
References Bamshad, M, T. Kivisild, et al. 2001 "Genetic Evidence on the Origins of Indian Caste Populations," Genome Research 11: 994-1004
Chaubey, G., Metspalu, M., Villems, M. R., Kivisild, T. 2007. Reply to Winters BioEssays 29(5): 499 Published Online: 20 Apr 2007 DOI: 10.1002/bies.20574.
Chaubey,G, Metspalu, M., Kivisild, T. and Villems, R. 2006 Peopling of South Asia: Investigating the Caste-Tribe Continuum in India, Bio Essays 29:91-100.
Chandrasekar, et al. 2009 Updating Phylogeny of Mitochondrial DNA Macrohaplogroup M in India: Dispersal of Modern Human in South Asian Corridor, PLoS ONE 4(10)e7447.
Kivisild, T. et al. 1999a Deep common ancestry of Indian and western-Eurasian mitochondrial DNA lineages, Current Biology 9:13311334. Kivisild, T. et al. 1999b The Place of the Indian mtDNA Variants in the Global Network of Maternal Lineages and the Peopling of the Old World, In Deka, R.P. ed. Genomic Diversity, 135-152 NY: S.S. Kluwer/Plenum Publishers.
Kivisild, T., et al. 2003 The Genetic Heritage of the Earliest Settlers Persists Both in Indian Tribal and Caste Populations, American Journal of Human Genetics 72(2): 313-332
Maca-Meyer, N., et al. 2001 Major Genomic Mitochondrial Lineages Delineate Early Human Expansions, BMC Genetics 2:13.
Metspalu, M., et al. 2004 More of the Extant mtDNA Boundaries in South and Southwest Asia were Likely Shaped During the Initial Settlement of Eurasia by Anatomically Modern Humans, BMC Genetics 5:26
Quintana-Murci, L., et al. 1999 "Genetic evidence of an early exit of Homo sapiens sapiens from Africa through eastern Africa," Nature Genetics 23: 437-441.
Rajkumar, R., et al. 2005 Phylogeny and Antiquity of M Macrohaplogroup Inferred from Complete mtDNA Sequence of Indian Specific Lineages, BMC Evolutionary Biology 5:26.
Sharma, G., et al. 2012 Genetic Affinities of the Central Indian Tribal Populations, PLoS ONE 7(2):e35246.
quote:Originally posted by Troll Patrol # Ish Gebor: Quetzalcoatl, is like. It's only peer reviewed if (whites) the western world has reviewed and consigned it.
It's somewhat the mindset of a colonist.
What Quetzalcoatl is saying is that western academia is superior, therefore credible.
Science is universal and does not have a color contingent. Scientists from all over the world (with all hues of skin color) publish in the mainline refereed journals. Here is a list of Nobel prize winners by country of birth http://www.nobelprize.org/nobel_prizes/lists/countries.html China (11) The Nobel Prize in Physics 1956 Walter H. Brattain "for their researches on semiconductors and their discovery of the transistor effect" The Nobel Prize in Physics 1957 Chen Ning Yang "for their penetrating investigation of the so-called parity laws which has led to important discoveries regarding the elementary particles" The Nobel Prize in Physics 1957 Tsung-Dao Lee "for their penetrating investigation of the so-called parity laws which has led to important discoveries regarding the elementary particles" The Nobel Prize in Physics 1998 Daniel C. Tsui "for their discovery of a new form of quantum fluid with fractionally charged excitations" The Nobel Prize in Physics 2009 Charles K. Kao "for groundbreaking achievements concerning the transmission of light in fibers for optical communication" The Nobel Prize in Chemistry 2010 Ei-ichi Negishi "for palladium-catalyzed cross couplings in organic synthesis"
Egypt The Nobel Prize in Chemistry 1999 Ahmed Zewail "for his studies of the transition states of chemical reactions using femtosecond spectroscopy"
Algeria The Nobel Prize in Chemistry 1999 Ahmed Zewail "for his studies of the transition states of chemical reactions using femtosecond spectroscopy"
India (7) The Nobel Prize in Physics 1930 Sir Venkata Raman "for his work on the scattering of light and for the discovery of the effect named after him" The Nobel Prize in Chemistry 2009 Venkatraman Ramakrishnan "for studies of the structure and function of the ribosome"
The Nobel Prize in Physiology or Medicine 1968 H. Gobind Khorana "for their interpretation of the genetic code and its function in protein synthesis" India (now Pakistan) (2) The Nobel Prize in Physics 1979 Abdus Salam "for their contributions to the theory of the unified weak and electromagnetic interaction between elementary particles, including, inter alia, the prediction of the weak neutral current" The Nobel Prize in Physics 1983 Subramanyan Chandrasekhar "for his theoretical studies of the physical processes of importance to the structure and evolution of the stars"
Japan (23) The Nobel Prize in Physics 1949 Hideki Yukawa "for his prediction of the existence of mesons on the basis of theoretical work on nuclear forces" The Nobel Prize in Physics 1965 Sin-Itiro Tomonaga "for their fundamental work in quantum electrodynamics, with deep-ploughing consequences for the physics of elementary particles" The Nobel Prize in Physics 1973 Leo Esaki "for their experimental discoveries regarding tunneling phenomena in semiconductors and superconductors, respectively" The Nobel Prize in Physics 2002 Masatoshi Koshiba "for pioneering contributions to astrophysics, in particular for the detection of cosmic neutrinos" The Nobel Prize in Physics 2008 Yoichiro Nambu "for the discovery of the mechanism of spontaneous broken symmetry in subatomic physics" The Nobel Prize in Physics 2008 Makoto Kobayashi "for the discovery of the origin of the broken symmetry which predicts the existence of at least three families of quarks in nature" The Nobel Prize in Physics 2008 Toshihide Maskawa "for the discovery of the origin of the broken symmetry which predicts the existence of at least three families of quarks in nature" The Nobel Prize in Physics 2014 Isamu Akasaki "for the invention of efficient blue light-emitting diodes which has enabled bright and energy-saving white light sources" The Nobel Prize in Physics 2014 Hiroshi Amano "for the invention of efficient blue light-emitting diodes which has enabled bright and energy-saving white light sources" The Nobel Prize in Physics 2014 Shuji Nakamura "for the invention of efficient blue light-emitting diodes which has enabled bright and energy-saving white light sources" The Nobel Prize in Physics 2015 Takaaki Kajita "for the discovery of neutrino oscillations, which shows that neutrinos have mass" The Nobel Prize in Chemistry 1981 Kenichi Fukui "for their theories, developed independently, concerning the course of chemical reactions" The Nobel Prize in Chemistry 2000 Hideki Shirakawa "for the discovery and development of conductive polymers" The Nobel Prize in Chemistry 2001 Ryoji Noyori "for their work on chirally catalysed hydrogenation reactions" The Nobel Prize in Chemistry 2002 Koichi Tanaka "for their development of soft desorption ionisation methods for mass spectrometric analyses of biological macromolecules" The Nobel Prize in Chemistry 2008 Osamu Shimomura "for the discovery and development of the green fluorescent protein, GFP" The Nobel Prize in Chemistry 2010 Akira Suzuki "for palladium-catalyzed cross couplings in organic synthesis" The Nobel Prize in Physiology or Medicine 1987 Susumu Tonegawa "for his discovery of the genetic principle for generation of antibody diversity" The Nobel Prize in Physiology or Medicine 2012 Shinya Yamanaka "for the discovery that mature cells can be reprogrammed to become pluripotent" The Nobel Prize in Physiology or Medicine 2015 Satoshi Ōmura "for their discoveries concerning a novel therapy against infections caused by roundworm parasites"
Mexico The Nobel Prize in Chemistry 1995 Mario J. Molina "for their work in atmospheric chemistry, particularly concerning the formation and decomposition of ozone"
Morocco The Nobel Prize in Physics 2012 Serge Haroche "for ground-breaking experimental methods that enable measuring and manipulation of individual quantum systems"
Taiwan The Nobel Prize in Chemistry 1986 Yuan T. Lee "for their contributions concerning the dynamics of chemical elementary processes"
Turkey The Nobel Prize in Chemistry 2015 Aziz Sancar "for mechanistic studies of DNA repair"
Venezuela The Nobel Prize in Physiology or Medicine 1980 Baruj Benacerraf "for their discoveries concerning genetically determined structures on the cell surface that regulate immunological reactions"
Posts: 833 | From: Austin, TX | Registered: Jan 2007
| IP: Logged |
quote:Originally posted by Troll Patrol # Ish Gebor: Quetzalcoatl, is like. It's only peer reviewed if (whites) the western world has reviewed and consigned it.
It's somewhat the mindset of a colonist.
What Quetzalcoatl is saying is that western academia is superior, therefore credible.
Science is universal and does not have a color contingent. Scientists from all over the world (with all hues of skin color) publish in the mainline refereed journals. Here is a list of Nobel prize winners by country of birth
I agree on that, but why did you judge those scientists from India and Pakisan?
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |
quote:Originally posted by Troll Patrol # Ish Gebor: @ Quetzalcoatl,
Are these the papers you are referring at, when you spoke of the M1-M3 transposition?
4. Kivisild T, Kaldma K, Metspalu M, Parik J, Papiha S, Villems R. 1999. The place of the Indian mtDNA variants in the global network of maternal lineages and the peopling of the Old World. In Deka R, Papiha SS. eds Genomic Diversity Kluwer Academic/Plenum Publishers New York p 135152.
5. Quintana-Murci L, Semino O, Bandelt H-J., Passarino G, McElreavey K, Santachiara-Benerecetti AS. 1999. Genetic evidence of an early exit of Homo sapiens sapiens from Africa through eastern Africa. Nat Genet 23: 437 441.
Yes, from my paper in Academia.edu
quote: . Kivisild (1999) and Quintana-Murci (1999) identified and named different haplotypes, one Indian and one African, as M1. This happens often and the usual procedure is for one of the authors to change the nomenclature. The final arbiter is www.phylotree.org that publishes an updated mtDNA tree. In this case, Quintana-Murcis (1999) identification of M1 was retained with haplotype markers 16129 16189 16249 16311. Kivisilds M1 had 16126 as the marker and this haplogroups name was changed to M3. The individuals in Kivisild (1999) labeled M1 did not disappear their designation changed to M3. Kivisilds compliance with the revised nomenclature is seen in Bamshad (2001). Numerous papers including some cited by Winters make it clear that there is no M1 in India although M3 with the 16126 marker (1999s M1) is definitely found (Bamshad 2001; Maca-Meyer 2001; Kivisild 2003- Dravidians; Metspalu 2004; Rajkumar 2005; Thangaraj 2006) Chandrasekar 2009; Shah 2012). In 2007, Winters (2007) repeated this distortion in a comment to a paper by Chaubey (2006). Chaubey, Metspalu, Kivisild, and Villems (2007) replied and clearly pointed out Winters misreporting of their paper. In 2008, Kivisild and Villems pointed this error to Winters by e-mail. Yet, he continues to use this argument and citation. This is academic dishonesty.
References Bamshad, M, T. Kivisild, et al. 2001 "Genetic Evidence on the Origins of Indian Caste Populations," Genome Research 11: 994-1004
Chaubey, G., Metspalu, M., Villems, M. R., Kivisild, T. 2007. Reply to Winters BioEssays 29(5): 499 Published Online: 20 Apr 2007 DOI: 10.1002/bies.20574.
Chaubey,G, Metspalu, M., Kivisild, T. and Villems, R. 2006 Peopling of South Asia: Investigating the Caste-Tribe Continuum in India, Bio Essays 29:91-100.
Chandrasekar, et al. 2009 Updating Phylogeny of Mitochondrial DNA Macrohaplogroup M in India: Dispersal of Modern Human in South Asian Corridor, PLoS ONE 4(10)e7447.
Kivisild, T. et al. 1999a Deep common ancestry of Indian and western-Eurasian mitochondrial DNA lineages, Current Biology 9:13311334. Kivisild, T. et al. 1999b The Place of the Indian mtDNA Variants in the Global Network of Maternal Lineages and the Peopling of the Old World, In Deka, R.P. ed. Genomic Diversity, 135-152 NY: S.S. Kluwer/Plenum Publishers.
Kivisild, T., et al. 2003 The Genetic Heritage of the Earliest Settlers Persists Both in Indian Tribal and Caste Populations, American Journal of Human Genetics 72(2): 313-332
Maca-Meyer, N., et al. 2001 Major Genomic Mitochondrial Lineages Delineate Early Human Expansions, BMC Genetics 2:13.
Metspalu, M., et al. 2004 More of the Extant mtDNA Boundaries in South and Southwest Asia were Likely Shaped During the Initial Settlement of Eurasia by Anatomically Modern Humans, BMC Genetics 5:26
Quintana-Murci, L., et al. 1999 "Genetic evidence of an early exit of Homo sapiens sapiens from Africa through eastern Africa," Nature Genetics 23: 437-441.
Rajkumar, R., et al. 2005 Phylogeny and Antiquity of M Macrohaplogroup Inferred from Complete mtDNA Sequence of Indian Specific Lineages, BMC Evolutionary Biology 5:26.
Sharma, G., et al. 2012 Genetic Affinities of the Central Indian Tribal Populations, PLoS ONE 7(2):e35246.
So, M1 became M3 somewhere in 1999.
The question I'm having here is, what is the "new" M = M1 reference?
Posts: 22234 | From: האם אינכם כילדי הכרית אלי בני ישראל | Registered: Nov 2010
| IP: Logged |





 UBBFriend: Email this page to someone!
UBBFriend: Email this page to someone!







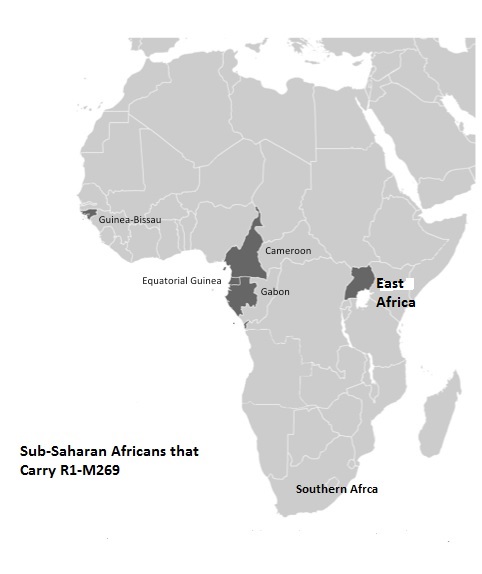

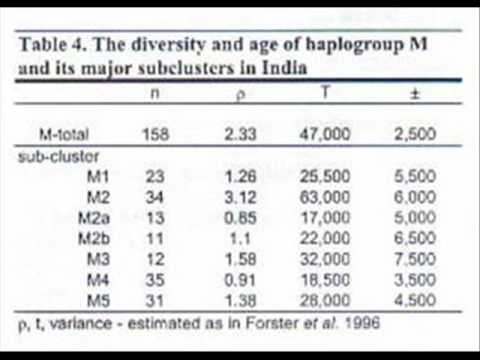







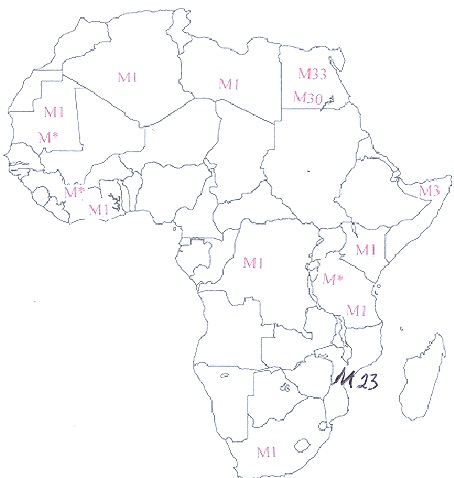
![[Frown]](frown.gif) Not again.
Not again. 
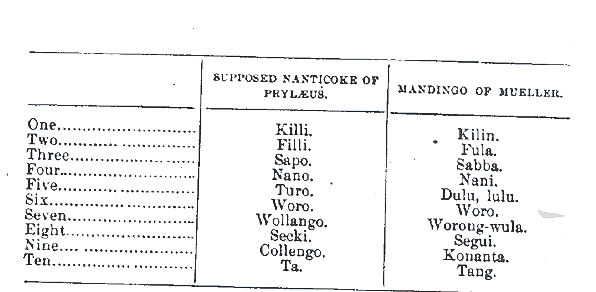




 Printer-friendly view of this topic
Printer-friendly view of this topic